3537
Axon diameter estimation in fixed human optic chiasm using diffusion weighted MR microscopy and microstructure-informed tractography1Department of Systems Neurosciences, University Medical Center Hamburg-Eppendorf, Hamburg, Germany, 2Department of Neurophysics, Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Germany, 3Berlin Ultrahigh Field Facility (B.U.F.F.), Max-Delbrueck-Center for Molecular Medicine in the Helmholtz Association, Berlin, Germany, 4Neurocomputation and Neuroimaging Unit, Department of Education and Psychology, Freie Universität Berlin, Berlin, Germany, 5Paul Flechsig Institute of Brain Research, University of Leipzig, Leipzig, Germany, 6Signal Processing Lab (LTS5), Ecole Polytechnique Federale de Lausanne, Lausanne, Switzerland, 7Department of computer science, University of Verona, Verona, Italy
Synopsis
We estimated mean axon diameter of ipsilateral bundles in a fixed human optic chiasm sample using diffusion weighted MR microscopy and COMMIT framework. We computed that the resolution limit for axon diameters of our dMRI data is about 1.3 µm, indicating that our approach is sensitive to a larger range of axon diameter distribution (ADD) than in-vivo dMRI studies. We found a similar pattern in the estimated ADD using MRI as was reported by a histological analysis of macaque monkeys’ optic chiasm, indicating that the estimated ADD provides similar sensitivity to regional differences in microstructure properties as in ex-vivo histology.
Introduction:
Biophysical models1 have been proposed to measure the axon diameter index (ADI) in white matter using diffusion MRI (dMRI). Recently, such a model for axon diameter estimation was incorporated into the COMMIT framework2, 3, 4 (Convex Optimization Modeling for Microstructure Informed Tractography). As opposed to the methods5, which work on a voxel-by-voxel basis, COMMIT estimates the axon diameter distribution (ADD) in the space of fiber pathways. One advantage of this approach is the improved comparability of MRI-derived ADDs with published histological data, which typically are determined for specific fiber pathways6, 7, 8. To achieve the necessary resolution limit for MRI-based ADD, this study used ultra-high signal-to-noise (SNR) ex-vivo dMRI microscopy in fixed human optic chiasm sample.Methods:
Sample: A human optic chiasm sample obtained at autopsy with prior informed consent (24 hrs postmortem, thrombosis) and approved by responsible authorities. Following standard Brain Bank procedures, blocks were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS, pH 7.4) at 4°C.
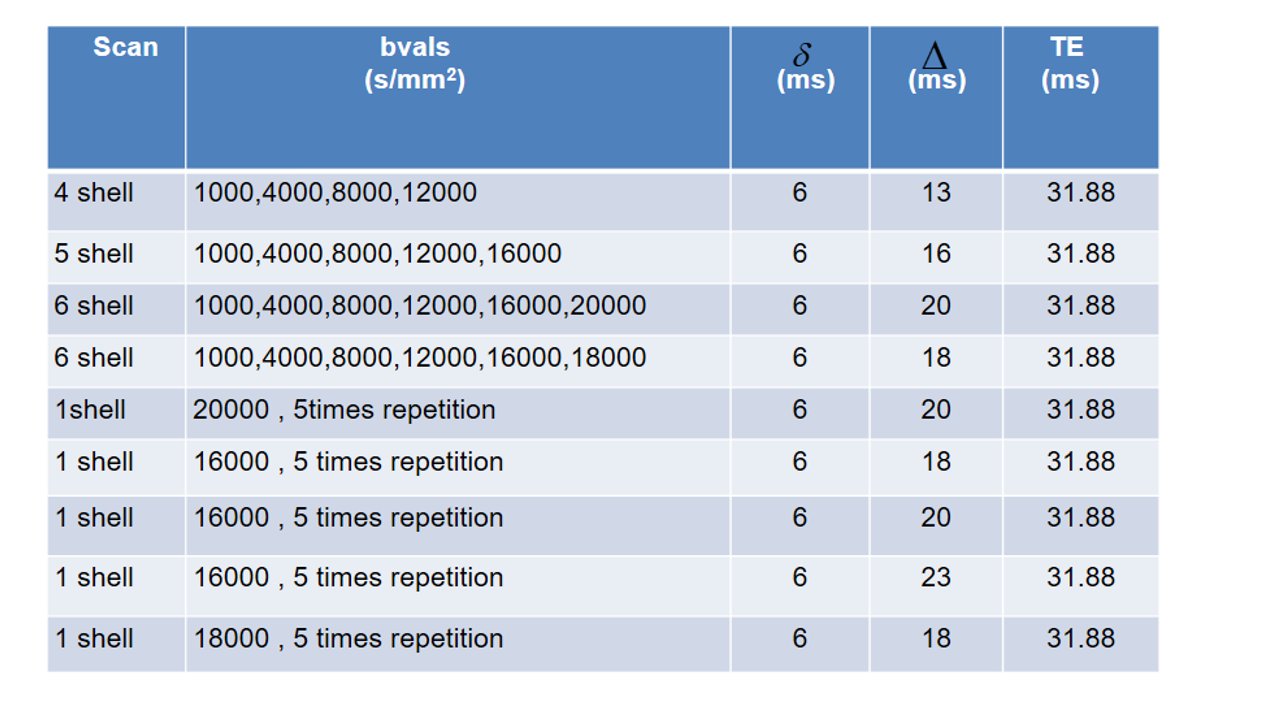
MRI: To facilitate ultra-high SNR, ex-vivo dMRI microscopy (details in Table 1), a small bore 9.4 T MR system (Bruker Biospin, Ettlingen, Germany) was used together with cryogenically-cooled TX/RX quadrature surface RF-coil and a high-performance gradient system (Gmax=738 mT/m).
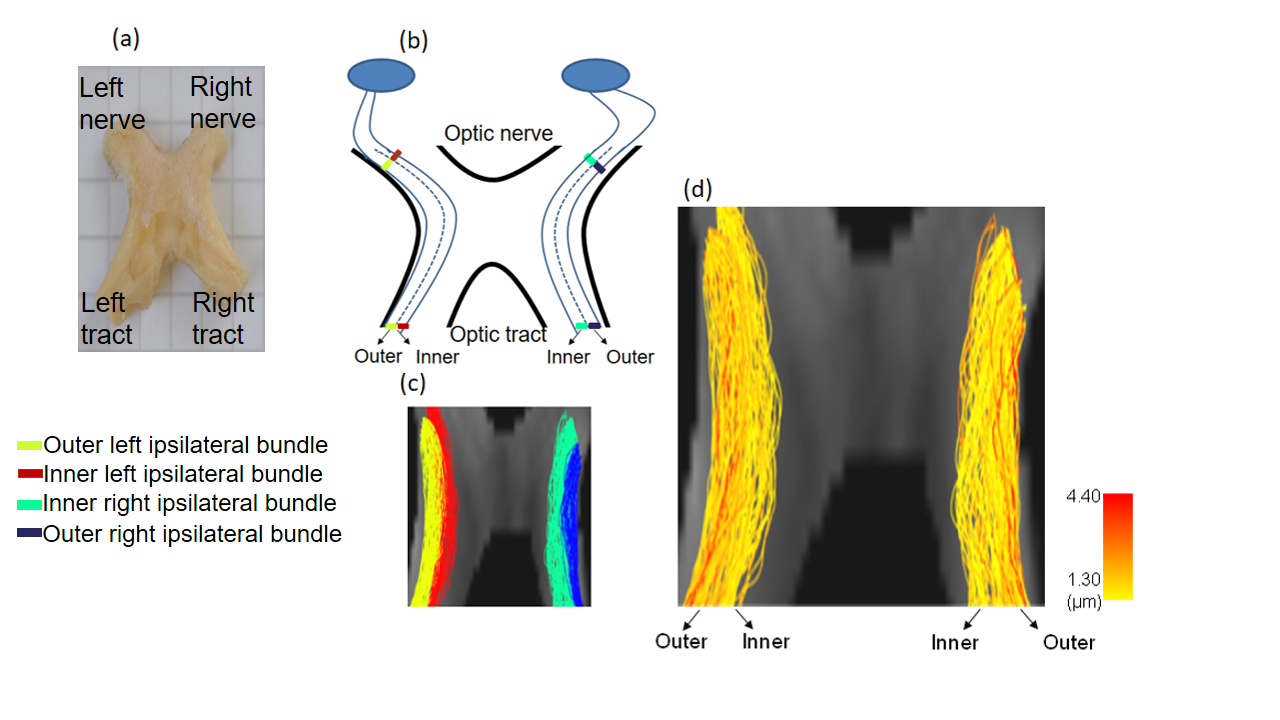
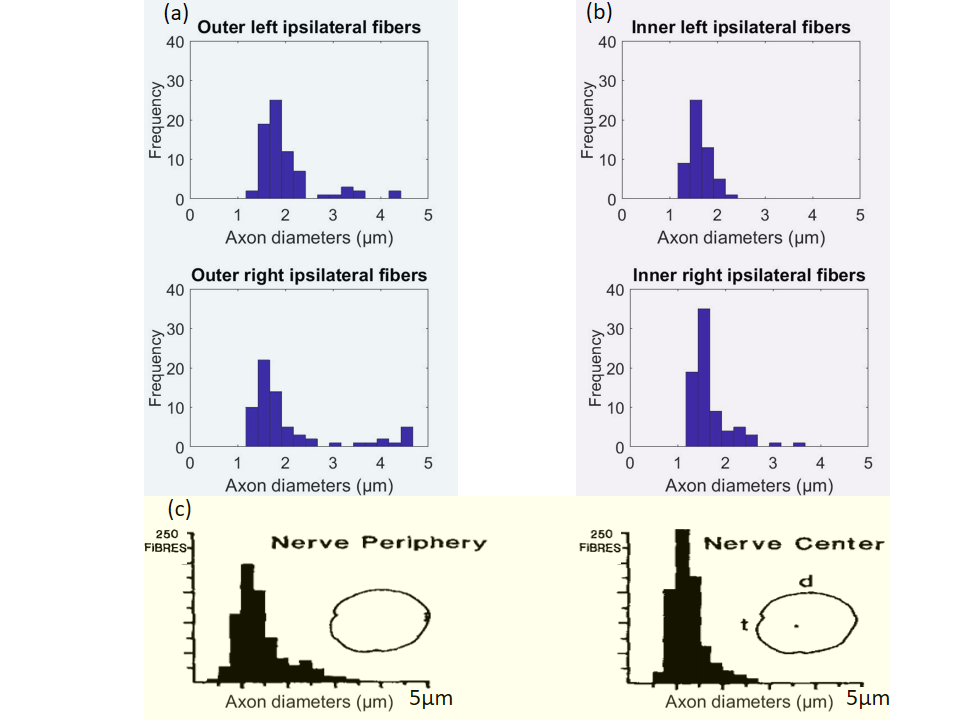
Tractography: To generate fiber bundles, we utilized the iFOD2 algorithm9 of MRtrix10. In this study, we restricted the analysis to the ipsilateral bundles of the optic chiasm7, 11 (Figure 3a-c), with the aim of comparing it to a published histological analysis8.
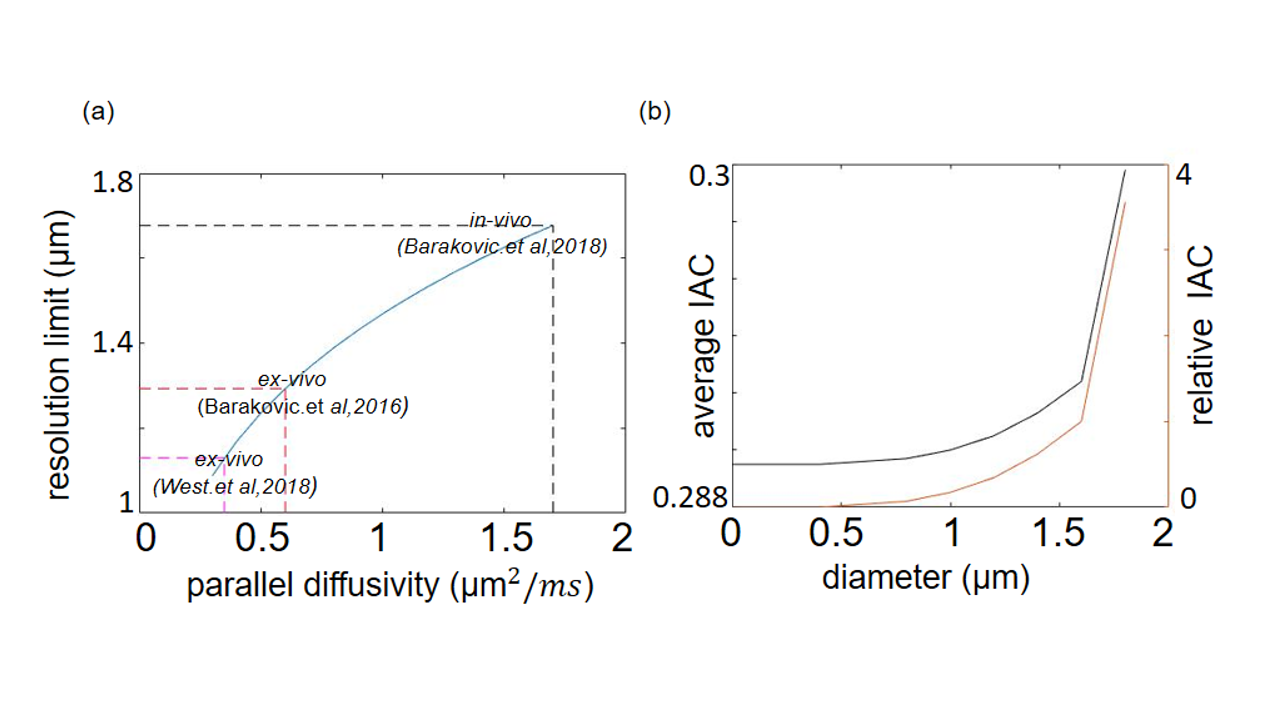
Microstructure analysis: This study used the Cylinder-Zeppelin-Ball (CZB) model12 in COMMIT and modeled the intra-axonal compartment (IAC) with 9 cylinders having diameters between 1.3 and 5.3 µm, the extra-axonal compartment with zeppelins having $$$d_{||}$$$=0.6 µm2/ms and three d⊥=[0.24, 0.36, 0.48] µm2/ms. The isotropic diffusion was modeled by balls with diso=2 µm2/ms. We estimated the resolution limit, the minimal resolvable axon diameter,of the acquisition protocol using two approaches:
(i) The analytical form, derived by 13 for the intra-cellular compartment assuming parallel cylinders:
$$$d_{min}=(\frac{768}{7}\frac{\bar{σ}D_{0}}{δγ^2g^2})^{\frac{1}{4}}$$$ (1)
For details see Figure 1.
(ii) An empirical experiment, where we characterized the MR signal using a set of CZB models, each of which consisting only of one cylinder with diameters (dim) between 0.4-1.8 µm, respectively. The aim of this experiment was to assess the relative change of the IAC estimated via the CZB model as compared to the IAC estimated by the Stick-Zeppelin-Ball (SZB) model12, which assumes a diameter of zero for all sticks, when cylinder’s diameters were increased.Hereby, we hypothesized that if the cylinder-diameter exceeds the resolution limit, more variance in the data will be explained as compared to the SZB model and thus the contribution of the IAC increases.
Results and Discussion:
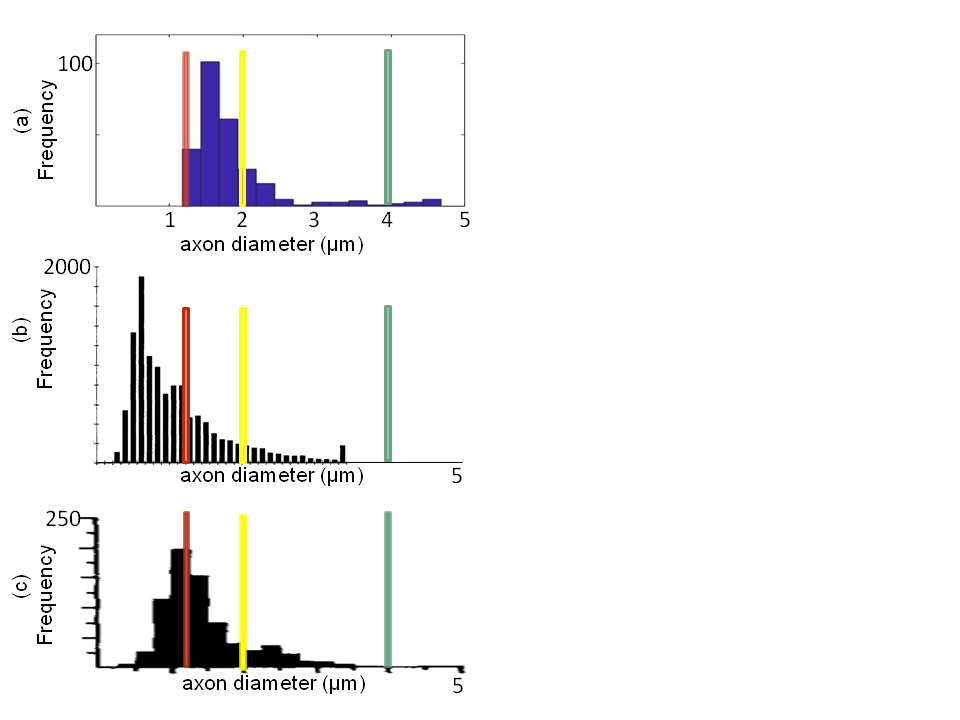
The theoretical resolution limit13 of MRI-based mean axon diameter estimation for fixed tissue was found to be 1.3 µm for our MRI protocol (Figure 1a). In an empirical experiment, we showed that above diameters of 0.4 µm the IAC estimated by the CZB model was larger than the IAC derived from the SZB model, suggesting that above this limit CZB explained more variance in the signal than the SZB model (Figure 1b). For the rest of the analyses, we used the theoretical resolution limit as this limit was supported by theory and experiment. Comparing the MRI-based ADD to the histology literature (Figure 2), we found that a large part of histological ADD could be recovered by using our protocol. The peak of the estimated ADD was overestimated as compared to the one reported for humans14, whereas it was comparable to the value reported for macaque monkeys8. The overestimation of the ADD could be related to neglecting axon undulation15 and/or fiber dispersion16. Finally, we found that the MRI-based ADD of the outer bundles (Figure 3d and 4a) showed a prominent tail towards larger diameters which was not visible for the ADD of the inner bundles (Figure 3d and 4b).Conclusion:
We showed that using ex-vivo dMRI microscopy and COMMIT, a large part of the histologically8, 14 reported ADD for the optic chiasm could be recovered, whereas in-vivo dMRI will likely be less efficient. Differences between ADD of inner and outer bundles of the optic chiasm reported in this work were similar to what is known from histology for macaque monkeys8, indicating that the proposed MR microscopy based ADD estimation provides similar sensitivity to regional differences in microstructure properties as ex-vivo histology. The latter conclusion has been drawn from an inter-species comparison, which has to be treated with caution. In future work, we will compare our results with ex-vivo histology data from a human chiasm sample.
Acknowledgements
This work was supported by the German Research Foundation (DFG Priority Program 2041 "Computational Connectomics”, [AL 1156/2-1;GE 2967/1-1; MO 2397/5-1; MO 2249/3–1], by the Emmy Noether Stipend: MO 2397/4-1) and by the BMBF (01EW1711A and B) in the framework of ERA-NET NEURON.References
1. Alexander, D.C. et al. Orientationally invariant indices of axon diameter and density from diffusion MRI. NeuroImage, 52(4):1374–1389, 2010.
2. Daducci, A. et al. COMMIT: Convex optimization modeling for microstructure informed tractography. IEEE transciption on medical imaging, 34(1), 246-257, 2015.
3. Baraković , M. et al. Assessment of bundle-specific axon diameter distributions using diffusion MRI tractography. OHBM conference, 2016.
4. Barakovic, M. et al. Assessing feasibility and reproducibility of a bundle-specific framework on in vivo axon diameter estimates at 300mT/m. ISMRM 2018.
5. Daducci, A. et al. Accelerated Microstructure Imaging via Convex Optimization (AMICO) from diffusion MRI data. Neuroimage, 105:32–44, 2015.
6. Liewald D.et al. Distribution of axon diameters in cortical white matter: an electron-microscopic study on three human brains and a macaque. Biol Cybern, 108:541–557, 2014.
7. Glen, J. et al. Segregated hemispheric pathways through the optic chiasm distinguish primates from rodents. Neuroscience 157 (3), 2008.
8. Reese, B.E.et al. Distribution of Axons According to diameter in the monkey's optic tract. Comparative neurology 260, 453-460, 1987.
9. Tournier, J-D.et al. Improved probabilistic streamlines tractography by 2nd order integration over fiber orientation distributions. Proceedings of the international society for magnetic resonance in medicine, 2010.
10. http://www.mrtrix.org
11. Richard S. Snell. Clinical Neuroanatomy. Lippincott Williams & Wilkins.2010.
12. Panagiotaki, E. et al.Compartment models of the diffusion MR signal in brain white matter: A taxonomy and comparison. NeuroImage, 59(3):2241–2254, 2012.
13. Nilsson, M.et al. Resolution limit of cylinder diameter estimation by diffusion MRI: The impact of gradient waveform and orientation dispersion. NMR in biomedicine 30(7), 2017.
14. Jonas JB et al.Histomorphometry of the Human Optic Nerve. Investigative Ophthalmology & Visual Science, 31.1990.
15. Ramirez-Manzanares, A. et al. Quantifying diameter overestimation of undulating axons from synthetic DW-MRI. ISMRM 2018.
16. Zhang H, et al. Axon diameter mapping in the presence of orientation dispersion with diffusion MRI. NeuroImage, 56(1): 1301-1315, 2011.
17. Constantidines, C. et al. Signal to noise measurement in magnitude image from NMR phased arrays. 1997.
18. Jones, D. et al. White matter integrity, fiber count, and other fallacies: The do's and don'ts of diffusion MRI. NeuroImage, 73:239-254, 2013.
Figures