3388
Characterization of the cerebro-cerebellar loop exploiting advanced tractography and dealing with thalamic synapsis1Neuroradiology Unit, Brain MRI 3T Research Center, IRCCS Mondino Foundation, Pavia, Italy, 2Department of Brain and Behavioural Sciences, University of Pavia, Pavia, Italy, 3School of Aerospace Mechanical and Mechatronic Engineering, University of Sydney, Sydney, Australia, 4Sydney Imaging, University of Sydney, Sydney, Australia, 5Brain Connectivity Center, IRCCS Mondino Foundation, Pavia, Italy, 6Queen Square MS Centre, Department of Neuroinflammation, UCL Queen Square Institute of Neurology, Faculty of Brain Sciences, University College London, London, United Kingdom, 7Brain MRI 3T Research Center, IRCCS Mondino Foundation, Pavia, Italy
Synopsis
Recently, advanced tractography has been used for assessing the feasibility of characterizing cerebro-cerebellar loop, composed of cerebello-thalamo-cortical and cortico-ponto-cerebellar pathways, acknowledging the issue of how tractography deals with polysynaptic connectivity, i.e. at thalamic level. In this work, polysynaptic cerebello-thalamo-cortical and cortico-ponto-cerebellar pathways were reconstructed using a multiplicative hypothesis for thalamic connectivity. Our findings revealed the importance of using such a multiplication factor for streamlines reaching the thalamic synapses to properly reconstruct cerebro-cerebellar connection. Furthermore, findings using polysynaptic tracts support the cerebellar role in cognition showing that cognitive/associative areas are the mainly involved in both the cerebello-thalamo-cortical and cortico-ponto-cerebellar pathways.
Introduction
The cerebellar involvement in cognition and motor functions is increasingly recognized(1). Cerebellar processing is mediated by the cerebro-cerebellar loop composed of: cerebello-thalamo-cortical (CTC) and cortico-ponto-cerebellar (CPC) pathways that have been previously reconstructed in humans in vivo using advanced MRI techniques(2,3). In particular, tractography algorithms that use anatomical constrains include deep grey matter masks to stop connectivity and reconstructing tracts in a mono-synaptic fashion(4,5). Hence, an open issue is how to reconstruct loops and connections including polysynaptic tracts, e.g. through the thalamus. A previous work(6) demonstrated that the specific hypothesis about synaptic connectivity at thalamic level affects tractography results. Hence, in the present work advanced tractography was used on high-quality data from the Human Connectome Project to achieve these aims: i) support the physiological evidence that the ratio of incoming to outgoing axons at thalamic level is comparable in humans to the range of 1:20 to 1:40 demonstrated in animals(7,8); ii) characterize the cerebro-cerebellar loop taking into account the polysynaptic tracts passing through the thalamus.Methods
Subjects: 28 subjects (10/18 male/female; 22-35 years) were acquired using a Siemens 3T Connectome Skyra scanner. Minimal pre-processed 3DT1-weighted images (1.25mm isotropic resolution) and high-quality diffusion-weighted data (1.25mm isotropic resolution, b=1000,2000,3000s/mm2, 90 directions/shell,18 b0 images) were downloaded from the ConnectomeDB (http://db.humanconnectome.org).
Whole-brain tractography and cerebro-cerebellar connections: These steps were conducted as previously described(6). Briefly, Multi-Shell-Multi-Tissue CSD technique (9), probabilistic streamline tractography (iFOD2)(10) and SIFT2(4) were used. Tractography was performed with 30 million streamlines and 4-classes segmentation within the Anatomically Constrained Tractography framework(5). After creation of an ad-hoc atlas(6) combining deep GM structures, AAL(11) and SUIT atlases(12), possible cerebro-cerebellar connections were selected from the whole-brain tractogram as those connecting cerebral cortical and subcortical structures with contralateral cerebellar cortical areas.
Polysynaptic CTC and CPC identification: From the cerebro-cerebellar connections selected in the previous step, CTC and CPC pathways were separated using the superior and middle cerebellar peduncle (SCP, MCP) masks as follows: i) right/left monosynaptic CTC pathways were identified selecting streamlines passing through the SCP and contralateral red nucleus masks and excluding streamlines passing through the other cerebellar peduncles. Then, polysynaptic CTC pathways were identified selecting streamlines entering/outing the thalamus in a one-to-all relationship(6); ii) right/left CPC pathways were identified selecting streamlines passing through the MCP mask and avoiding streamlines passing through the contralateral MCP mask.
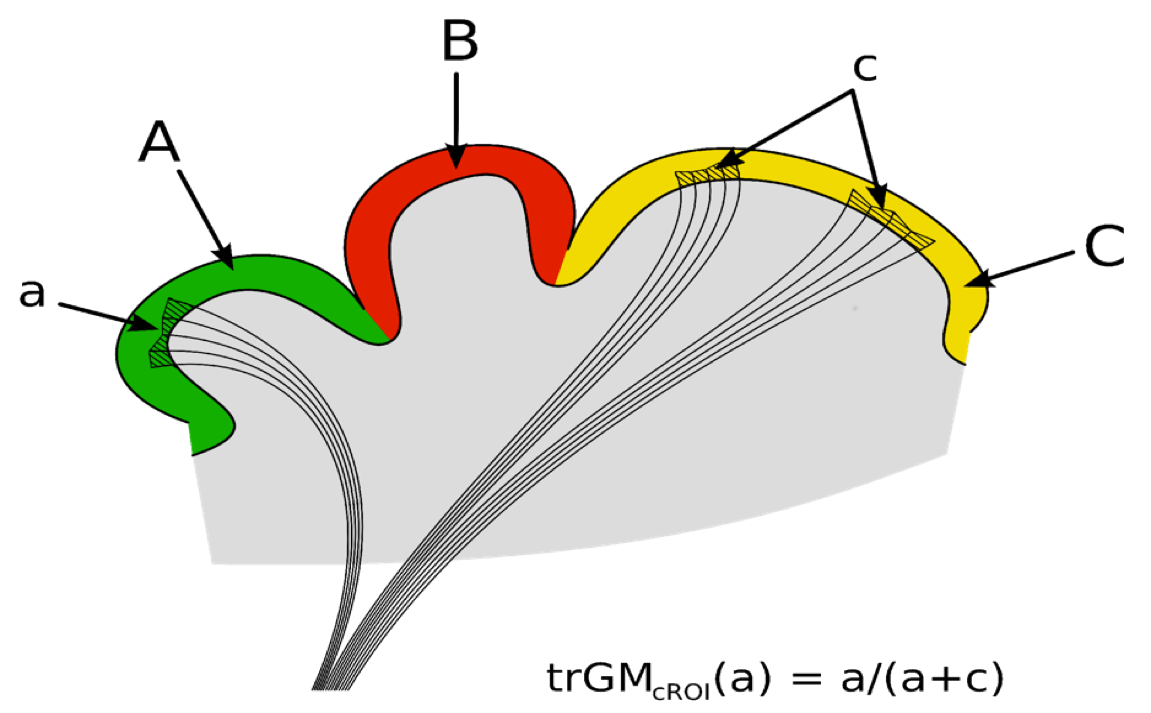
For each CTC/CPC pathway, the proportion of streamlines projecting to a particular cortical ROI was calculated by assessing the trGMcROI index (Figure1)(2).
Connectivity matrices creation: To characterize CTC and CPC pathways, connectivity matrices were created using the weigh-sum of streamlines as edges and cortical/subcortical regions as nodes.
Result
The reconstruction of polysynaptic CTC pathways revealed that the ratio between incoming and outgoing thalamic streamlines was in the range of 1:36 to 1:165 with a mean of 1:(71±26) across all subjects.
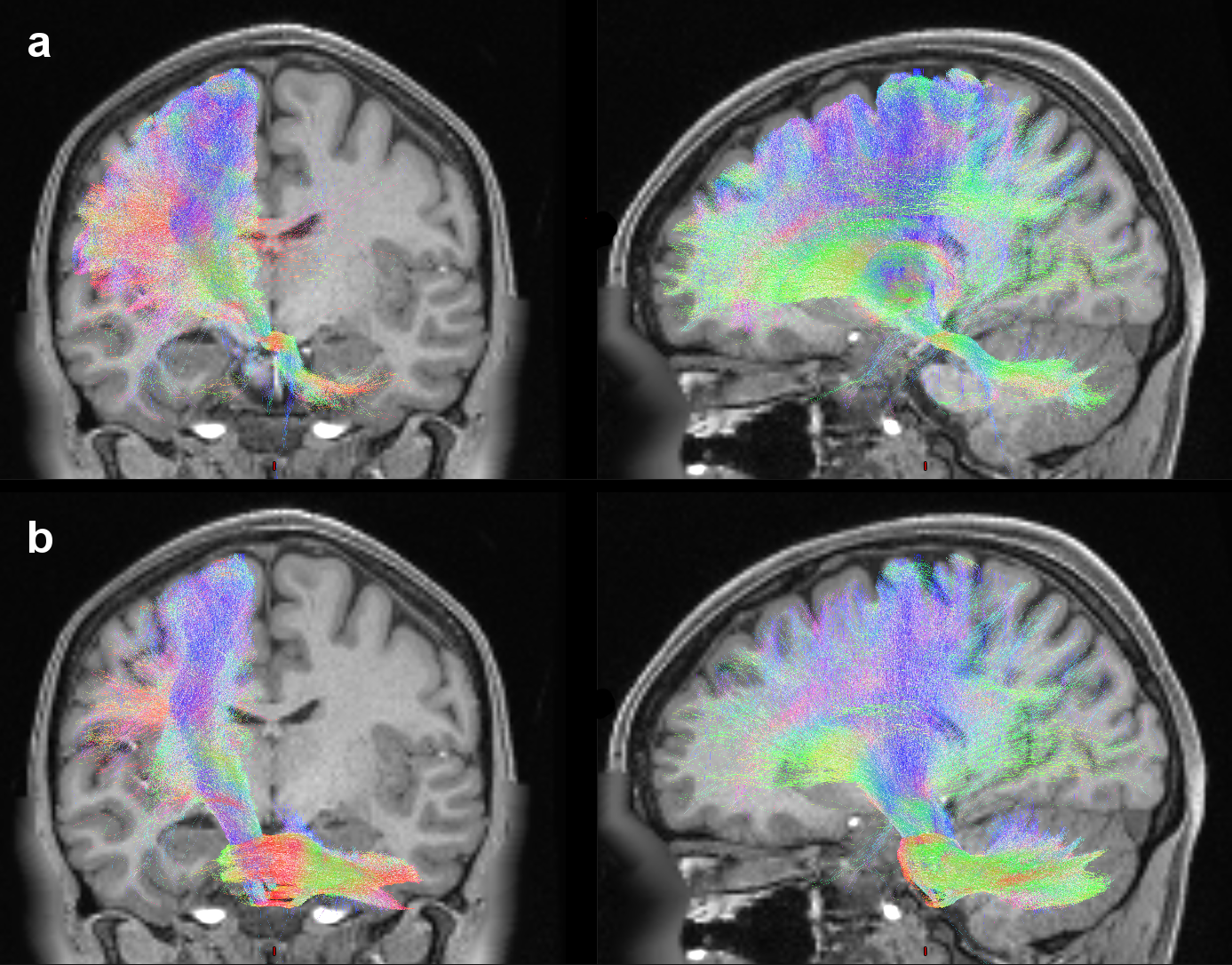
Figure2 shows both the polysynaptic CTC pathway, reconstructed adding the thalamic streamlines to the monosynaptic CTC ones (a), and the CPC pathway (b).
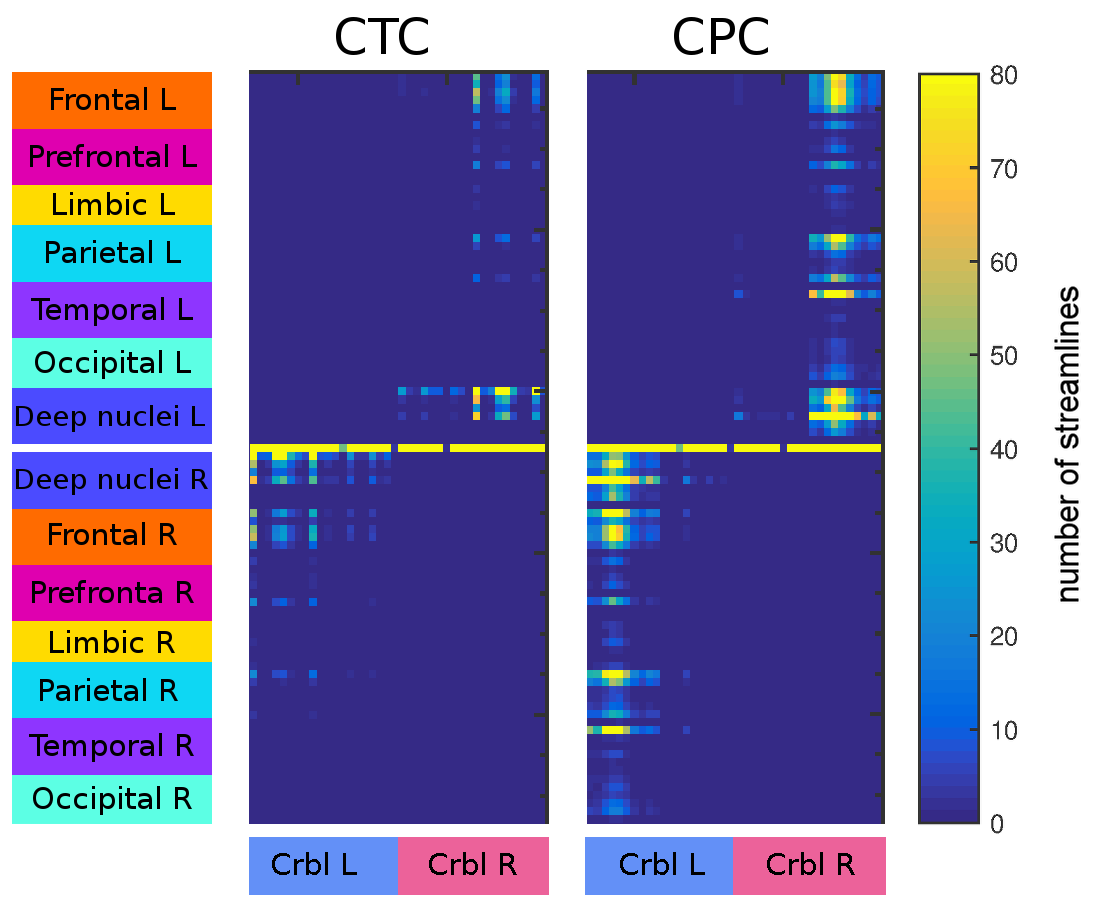
Figure3 shows connectivity matrices of the polysynaptic CTC and CPC pathways revealing that CPC pathways connect the cerebellum with a higher number of cerebral regions and, moreover, that strengths of CPC connections are generally higher with respect to those of CTC connections.
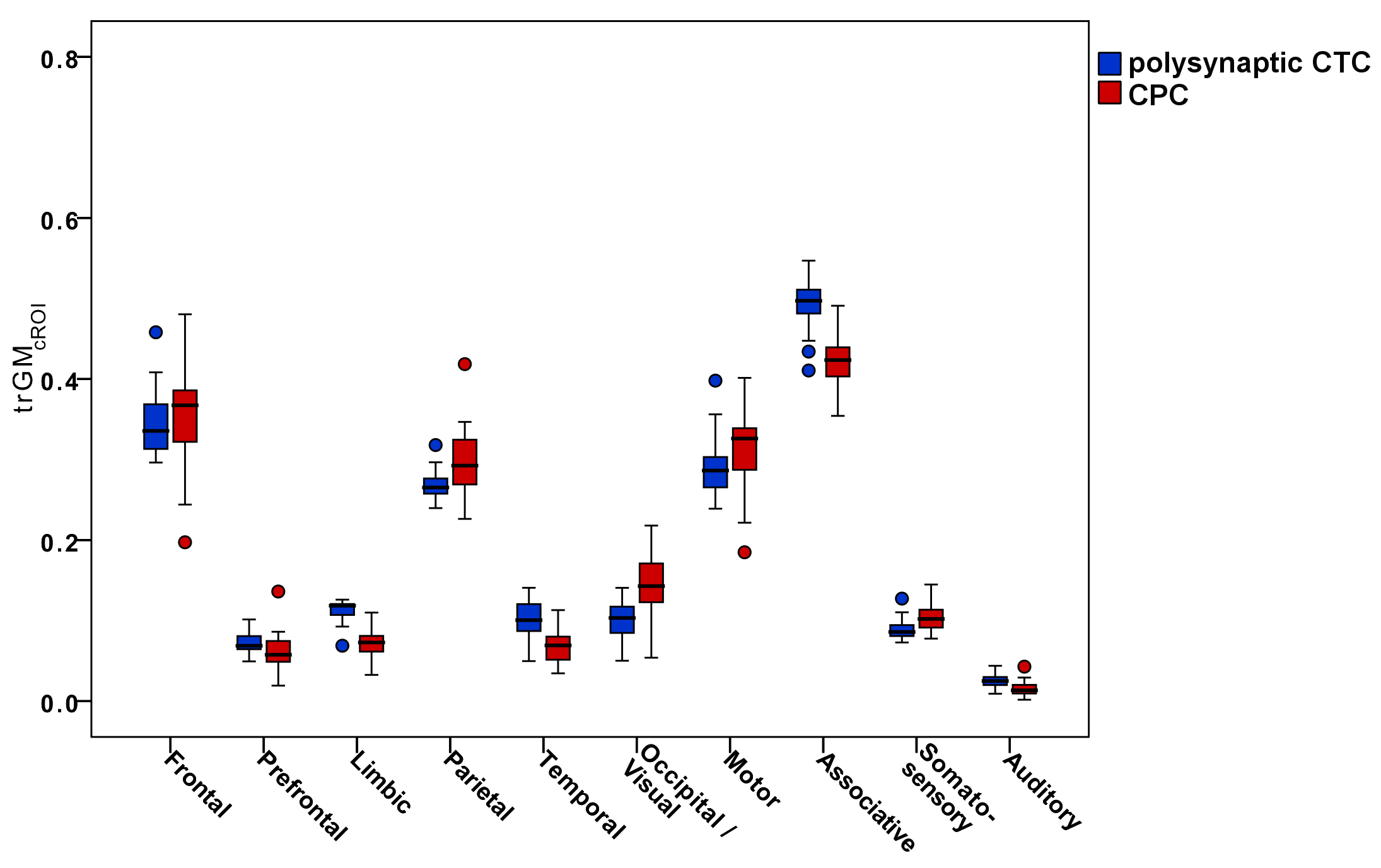
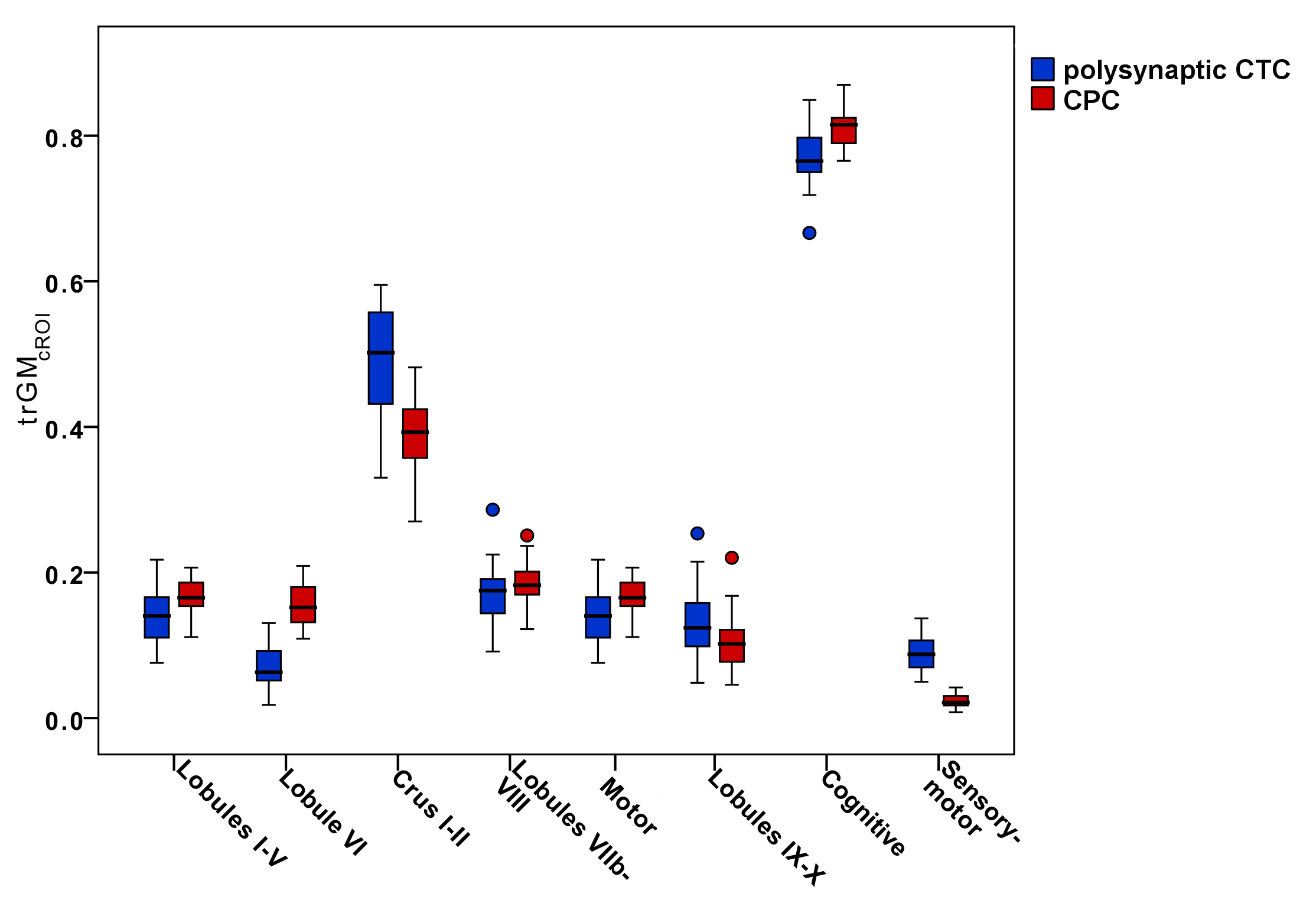
Figure4-5 report boxplots of the trGMcROI values for CTC and CPC pathways with respect to cerebral and cerebellar parcellations, respectively.
Discussion
The first important finding of our work is that, using the one-to-all relationship for streamlines at thalamic synaptic level, the mean ratio between incoming/outgoing thalamic streamlines seems to agree with previous physiological studies in animals, which assert that synapses amplify the number of entering connections with a multiplication factor between 20 and 40(7,8).
Furthermore, our results confirm that cerebro-cerebellar connections could operate in a closed loop(1,3) interacting with several cortical regions. Polysynaptic CTC pathways connect several areas of the cerebellum with the frontal cortex, but also with prefrontal, parietal and temporal cortices, while CPC pathways mainly connect several areas of the cerebral cortex with CrusI-II of the cerebellum. Furthermore, the majority of both CTC and CPC streamlines projects to cognitive/associative areas.
In conclusion, our work has highlighted the importance of using a multiplication factor for streamlines reaching the thalamic synapses when reconstructing axonal bundles in vivo and support the critical role of the cerebellum both in motor and executive control using state-of-the-art technology, i.e. high-resolution data and advanced analyses. Nevertheless, further multi-level studies are warranted to demonstrate the actual multiplication factor in human and propose algorithms able to solve this major issue in order to give proper quantitative characterization of the tracts involved in the cerebro-cerebellar loop.
Acknowledgements
Data were provided by the Human Connectome Project, WU-Minn Consortium (Principa lInvestigators: David Van Essen and Kamil Ugurbil; 1U54MH091657) funded by the 16 NIH Institutes and Centers that support the NIH Blueprint for Neuroscience Research; and by the McDonnell Center for Systems Neuroscience at Washington University.
University of Pavia, IRCCS Mondino Foundation (Pavia, Italy), UCL-UCLH Biomedical Research Centre (London, UK), National Health and Medical Research Council of Australia, the Australian Research Council for funding and for ongoing support.
References
1. D'Angelo E, Casali S. Seeking a unified framework for cerebellar function and dysfunction: from circuit operations to cognition. Front Neural Circuits (2013) 6(116):1-23. doi: 10.3389/fncir.2012.00116
2. Palesi F, Tournier J-D, Calamante F, Muhlert N, Castellazzi G, Chard D, et al. Contralateral cerebello-thalamo-cortical pathways with prominent involvement of associative areas in humans in vivo. Brain Struct Funct (2015) 220(6):3369-84. doi: 10.1007/s00429-014-0861-2.
3. Palesi F, Rinaldis AD, Castellazzi G, Calamante F, Muhlert N, Chard D, et al. Contralateral cortico-ponto-cerebellar pathways reconstruction in humans in vivo: implications for reciprocal cerebro-cerebellar structural connectivity in motor and non-motor areas. Sci Rep (2017) 7(1). doi: 10.1038/s41598-017-13079-8.
4. Smith RE, Tournier J-D, Calamante F, Connelly A. SIFT2: Enabling dense quantitative assessment of brain white matter connectivity using streamlines tractography. NeuroImage (2015) 119:338-51. doi: 10.1016/j.neuroimage.2015.06.092.
5. Smith RE, Tournier J-D, Calamante F, Connelly A. Anatomically-constrained tractography: improved diffusion MRI streamlines tractography through effective use of anatomical information. NeuroImage (2012) 62(3):1924-38. doi: 10.1016/j.neuroimage.2012.06.005.
6. Palesi F, Calamante F, Savini G, D'Angelo E, Gandini Wheeler-Kingshott CAM. The unresolved problem of synaptic connectivity in the context of the cerebro-cerebellar loop. Proc Intl Soc Mag Reson Med (2018) 26
7. Aumand TD, Horner MK. Ramification and Termination of Single Axons in the Cerebellothalamic Pathway of the Rat. The journal of comparative neurology 376:420-430 (1996).
8. Mason A, Ilinsky IA, Maldonado S, Kultas-Ilinsky K. Thalamic terminal fields of individual axons from the ventral part of the dentate nucleus of the cerebellum in Macaca mulatta.The journal of comparative neurology( 2000) Jun 5;421(3):412-28.
9. Jeurissen B, Tournier JD, Dhollander T, Connelly A, Sijbers J. Multi-tissue constrained spherical deconvolution for improved analysis of multi-shell diffusion MRI data. NeuroImage (2014) 103:411-26. doi: 10.1016/j.neuroimage.2014.07.061.
10. Tournier J-D, Calamante F, Connelly A, editors. Improved probabilistic streamlines tractography by 2 nd order integration over fibre orientation distributions. Proc Intl Soc Mag Reson Med (2010) 18.
11. Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, Delcroix N, et al. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage (2002) 15(1):273-89. doi: 10.1006/nimg.2001.0978.
12. Diedrichsen J, Balsters JH, Flavell J, Cussans E, Ramnani N. A probabilistic MR atlas of the human cerebellum. NeuroImage (2009) 46(1):39-46. doi: 10.1016/j.neuroimage.2009.01.045.
Figures