3371
Reducing false-positive connections using hierarchical microstructure-informed tractography1Computer Science Department, University of Verona, Verona, Italy, 2Athena Project-Team, Inria Sophia-Antipolis Méditerranée, Université Côte d'Azur, Nice, France, 3Signal Processing Laboratory 5 (LTS5), École Polytechnique Fédérale de Lausanne, Lausanne, Switzerland, 4Radiology Department, University Hospital Center (CHUV) and University of Lausanne (UNIL), Lausanne, Switzerland, 5Sherbrooke Connectivity Imaging Laboratory (SCIL), University of Sherbrooke, Sherbrooke, QC, Canada, 6Department of Nuclear Medicine and Radiobiology, Faculty of Medicine and Health Science, Sherbrooke Molecular Imaging Center, Sherbrooke, QC, Canada
Synopsis
Tractography has proven particularly effective for studying non-invasively the neuronal architecture of the brain, but recent studies have showed that the high incidence of false-positives can significantly bias any connectivity analysis. Last year we presented a method that extended COMMIT framework to consider the prior knowledge that white matter fibers are organized in bundles. Inspired by this, here we propose another extension to further improve the quality of the tractography reconstructions. We introduce a novel regularization term based on the multilevel hierarchy organization of the human brain and we test the results on both synthetic phantom and in vivo data.
Introduction
Tractography employs diffusion MRI (dMRI) data to non-invasively reconstruct the trajectories of major white-matter tracts1. Thanks to this unique ability it has been widely used to study the structural brain connectivity. However, recent studies have shown that the accuracy of the reconstructions is inherently limited2: existing algorithms suffer from an intrinsic trade-off between sensitivity, i.e. false-negatives connections (FNC), and specificity, i.e. false-positives (FPC), where specificity seems to be the major bottleneck3 when studying the topological properties of brain networks4.
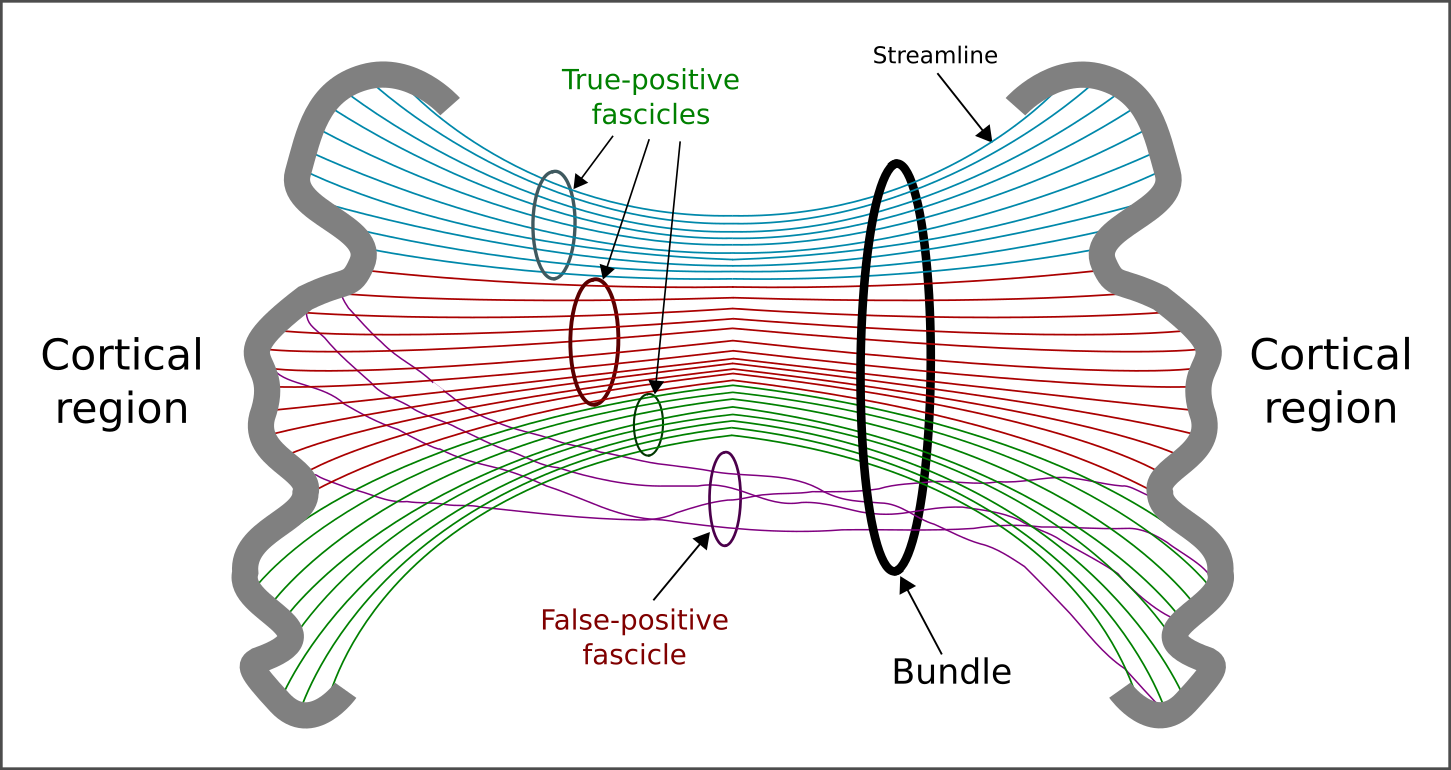
To overcome this limitation, a number of solutions have been proposed5. A common approach consists of filtering the reconstructed fibers (or streamlines) by using forward models and assessing their contribution using optimization. The Convex Optimization Modeling for Microstructure Informed Tractography6-7 (COMMIT) is one of them. Last year we extended it by adding a regularization term which allowed us to inject into COMMIT prior knowledge on the anatomical organization of the brain8 and which has proved very effective in reducing false positives8. A natural extension of this formulation is to further refine this prior by considering that the human brain has a hierarchical organization9, i.e. axonal bundles connecting different cortical regions may be composed by sub-bundles (or fascicles). Here, we propose a novel regularization term based on a multilevel-hierarchical organization of the streamlines and we test the benefits of considering such anatomical prior in COMMIT using synthetic and in vivo data.
Methods
Proposed approach. The observation model is $$$\mathbf{y}=\mathbf{Ax}+\eta$$$, where the matrix $$$\mathbf{A}$$$ implements a generic multi-compartment model to characterize the white-matter (WM) tissue, $$$\mathbf{x}$$$ are the contributions of the model’s compartments used for explaining the dMRI data $$$\mathbf{y}$$$, and $$$\eta$$$ represent the noise.
To consider the prior knowledge that WM fibers are organized in bundles, the problem is solved with non-negative least squares (NNLS) coupled with a regularization term that penalizes or promotes groups of streamlines connecting different pairs of cortical regions. The mathematical formulation is then $$$\text{argmin}_{\mathbf{x}\geq0}||\mathbf{Ax}-\mathbf{y}||^2_2+\lambda\sum_{g \in G}||\mathbf{x}^{(g)}||_2$$$, where $$$G$$$ represents a partition in groups of the streamlines in the tractogram. This formulation was presented in 8 and we call it gNNLS.
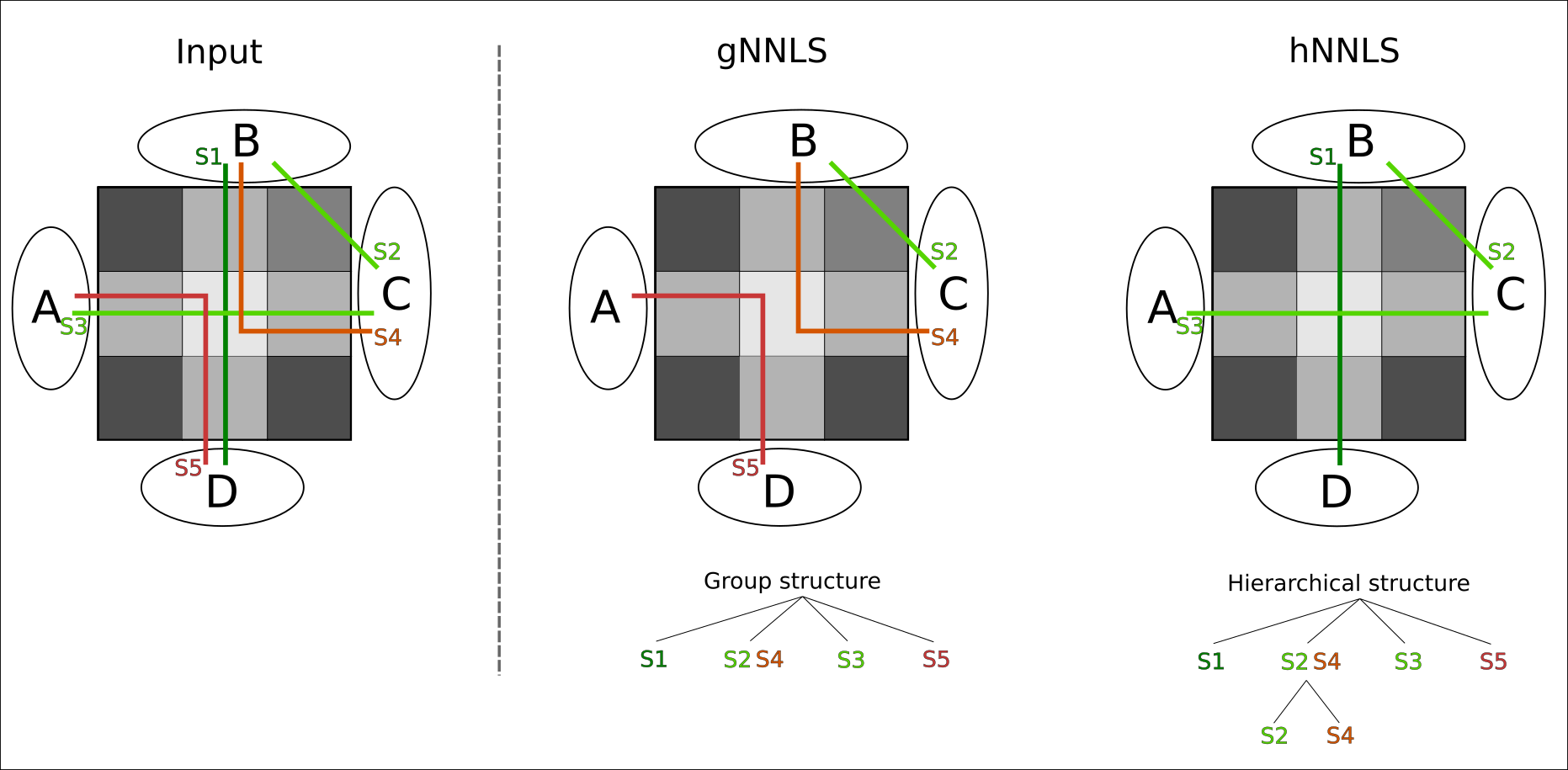
To further refine this prior and consider that axons may have a hierarchical organization9, we redefine the partition $$$G$$$ by creating a multilevel-hierarchical structure of the streamlines (Fig.1). The first level uses the same partitioning of gNNLS; then, for each a group we create a second level by clustering the streamlines of each group using QuickBundle10. This new partitioning may allow filtering out those false-positives fascicles that are part of true-positive bundles (violet in Fig.1), which gNNLS would not be able to distinguish. We call this formulation hNNLS.
For sake of simplicity, in both formulations we implemented a simple forward-model which assigns a signal contribution to each streamline that is proportional to its length inside a voxel. The total amount of streamlines that traverse a voxel must sum to the voxel's total intra-axonal signal fraction. This value can be estimated using standard models such as NODDI11 and SMT12; in this work we used the latter.
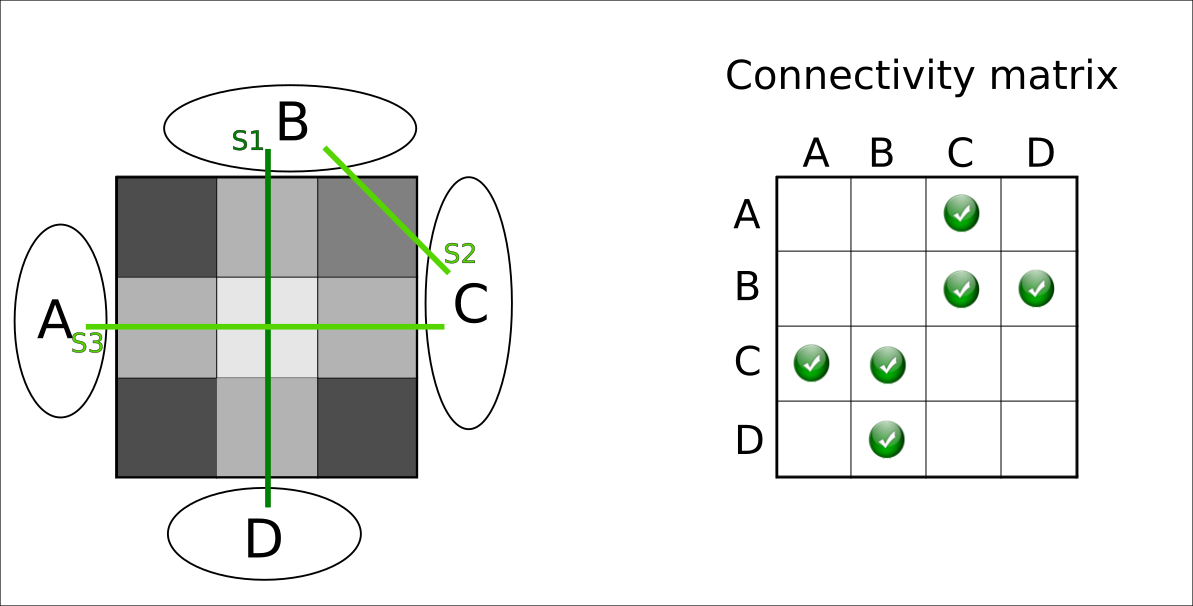
Experimental settings. We tested gNNLS and hNNLS on both synthetic and in-vivo data. The synthetic phantom is illustrated in Fig.2 and is used for illustrative purposes. In vivo data was acquired on a Siemens Prisma 3T scanner using 5 b-values in the range [300,3000]s/mm2, and was used to reconstruct 10M streamlines with iFOD213. A T1-MPRAGE image was acquired to perform gray-matter parcellation using FreeSurfer.
Results and Discussion
Although gNNLS was shown to be very effective for removing FPC8, in the ambiguous configuration of Fig.2 it fails to recover the ground-truth configuration, while hNNLS successfully estimates it (Fig.3). To keep the streamline S2 (true-positive) between B-C, which is required to explain the signal in the upper-right voxel, gNNLS has to keep as well S4, which is a false-positive streamline but belongs to the same group of S2. As a consequence, also S1 is removed and, in turn, S3. Because of its finer partition, hNNLS is instead able to decouple their contributions.
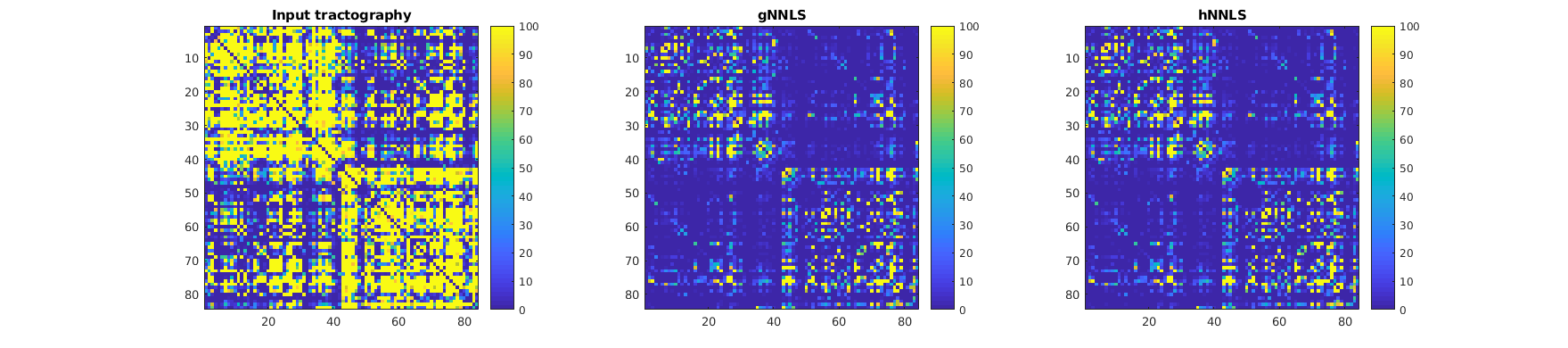
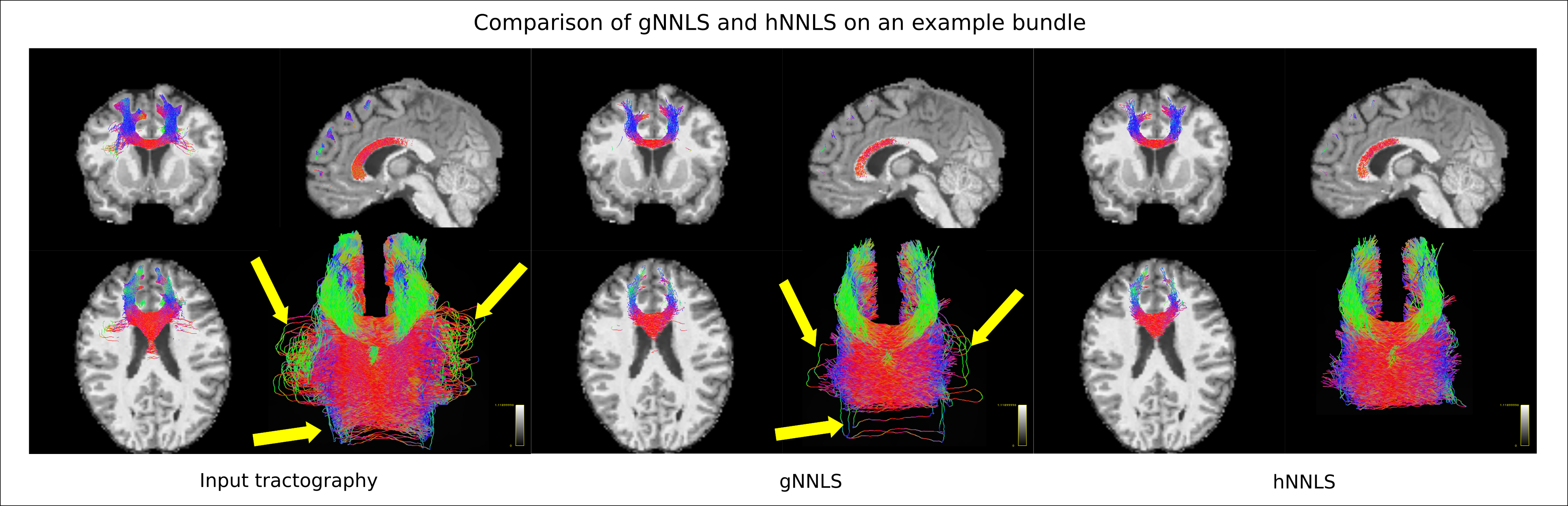
In in vivo data, a qualitative look at the connectivity matrices shows that the number of streamlines kept by gNNLS and hNNLS is similar (Fig.4). Yet, by visually inspecting known bundles we can appreciate the different filtering of the two formulations (Fig.5). Indeed, hNNLS is able to filter out streamlines that follow a strange path by keeping intact the structure of the bundle.
Conclusion
We showed that by adding to COMMIT a regularization term to promote a hierarchical structure of the streamlines, it is possible to further improve the quality of tractography reconstructions. We speculate that this improved reconstruction accuracy can have a significant impact on any analysis of structural brain connectivity.
Acknowledgements
This work was supported by the Rita Levi Montalcini Programme of the Italian Ministry of Education, University and Research (MIUR), as well as the Faculté de biologie et de médecine of the Lausanne University Hospital Center (CHUV).References
- Basser P, Pajevic S, Pierpaoli C, Duda J, Aldroubi A. In Vivo
Fiber Tractography Using DT-MRI Data. Magn. Reson. Med.
2000; 44:625– 632.
- Thomas C, Ye FQ, Irfanoglu MO, Modi P, Saleem KS, Leopold DA,Pierpaoli C. Anatomical accuracy of brain connections derived from diffusion MRI tractography is inherently limited. Proc Natl Acad Sci USA. 2014; 111(46):16574-9
- Maier-Hein KH et al. The challenge of mapping the human connectome based on diffusion tractography. Nature Communications. 2017; 8:1349
- Zalesky A, Fornito A, Cocchi L, Gollo LL, van den Heuvel MP, Breakspear M. Connectome sensitivity or specificity: which is more important? Neuroimage. 2016; 142:407-420
- Daducci A, Dal Palù A, Descoteaux M, Thiran JP. Microstructure Informed Tractography: Pitfalls and Open Challenges. Front Neurosci. 2016; 10:247
- Daducci A, Dal Palù A, Lemkaddem A, Thiran JP. A convex optimization framework for global tractography. In Proc. IEEE ISBI. 2013; 524–7
- Daducci A, Dal Palù A, Lemkaddem A, Thiran JP. COMMIT: Convex Optimization Modeling for Microstructure Informed Tractography. IEEE Trans Med Imaging. 2014; 33(1):246–57
- Daducci A, Barakovic M, Girard G, Descoteaux M, Thiran JP. Reducing
false positives in tractography with microstructural and anatomical
priors. Proc. Int. Soc. Magn. Reson. Med. 2018; 26, 0038
- Hagmann P. From diffusion MRI to brain connectomics. 2005; PhD thesis (chapter 10).
- Garyfallidis E, Brett M, Correia MM, Williams GB, Nimmo-Smith I. QuickBundles, a method for tractography simplification. Front Neurosci. 2012; 6 (175).
- Zhang H, Schneider T, Wheeler-Kingshott CA, Alexander DC. NODDI: practical in vivo neurite orientation dispersion and density imaging ofthe human brain. Neuroimage. 2012; 61(4):1000-16
- Kaden E, Kelm ND, Carson RP, Does MD, Alexander DC.
Multi-compartment microscopic diffusion imaging. Neuroimage. 2016;
139:346-359
- Tournier JD, Calamante F, Connelly A. Improved probabilistic streamlines tractography by 2nd order integration over fibre orientation distributions. Proc. Int. Soc. Magn. Reson. Med. 2010, 1670
Figures