3321
Progesterone, not estrogen modulates cerebral blood flow across the menstrual cycle1Médecine nucléaire et radiobiologie, Université de Sherbrooke, Sherbrooke, QC, Canada, 2Psychology, Bishop's University, Sherbrooke, QC, Canada, 3Pediatrie, Université Sherbrooke, Sherbrooke, QC, Canada, 4Radiologie diagnostic, Université de Sherbrooke, Sherbrooke, QC, Canada
Synopsis
The sex hormones estrogen (EST) and progesterone (PROG) are known vasodilators, yet little is known about how large changes in concentrations across the menstrual cycle (MC) influences CBF. This study aimed to determine how fluctuations in EST and PROG across the MC influence CBF and cerebral arteries. CBF and arteries were evaluated twice in female participants when PROG was low and high. There were region-specific and dose-dependent effects of PROG (but not EST) on CBF, but not arterial diameters. This indicates that cerebral vascular function in women is dynamic across the MC and is not due to vasodilatory effects of EST or PROG.
Introduction
Despite minimal differences in conventional magnetic resonance imaging (MRI) measures of brain morphology, men and women show significant differences in cerebral blood flow (CBF)1,2. One explanation for this is that the female brain is subject to a different hormonal environment3. For instance, estrogen (EST) and progesterone (PROG) levels are not only higher in women, but also dramatically change across the ~28-day menstrual cycle (MC)4.
Both EST and PROG are known vasodilators5,6, suggesting that cerebral vascular structure and function is not only different across the sexes but may also vary within the same women during the MC. As it stands, no study has yet investigated how changes in circulating hormone levels affect the size of blood vessels or CBF in the female brain. Therefore, the objective of this study was to determine how MC-related fluctuations in EST and PROG alter cerebral arterial diameters and CBF.
Methods
We evaluated 11 naturally cycling women not taking any form of hormonal contraception in the follicular (low serum PROG, low serum EST) and luteal phase (high serum PROG, high serum EST) of their MC and two women on combined oral contraception (i.e., no changes in PROG or EST) on week two and three of the pill packet.
Regional CBF (rCBF) was measured using pCASL (background suppression, label duration=1650ms, postlabel delay=1800ms, 2D multislice EPI readout, TR/TE 4246/16ms, 22 4mm, 3x3 resolution, FOV=240X240mm, effective temporal resolution 8492ms) and flow was determined using the FSL BASIL toolbox7. Arterial morphology was obtained using Time of Flight (TOF) magnetic resonance angiography (MRA) consisting of one high-resolution TOF MRA with only one chunk (FOV ¼ 200mmX200X30mm, TR ¼ 23ms, TE ¼ 3.6ms, FA ¼ 180 , parallel imaging (SENSE) acceleration factor ¼ 2.8, acquisition resolution of 0.4X0.4X0.8mm for a 0.4mm isotropic reconstructed resolution, acquisition time of 2min34s) centered on the circle of willis acquired as a slab image providing high resolution image of major supply vessels in the brain (figure 1b). An in-house method was used to isolate arterial vessels and extract diameters8,9. Following the MRI protocol, blood tests were performed to determine serum EST and PROG. A whole brain, voxel-wise correlation (p<0.01, cluster > 1000 voxels for cortex and cluster > 150 voxels for the midbrain) was performed with the MC-change in serum EST and PROG (ΔEST, ΔPROG) and change in rCBF (ΔrCBF), along with the diameters of the basilar artery (ΔBA).
Results
As expected, ΔPROG was highly variable across women (Figure 1a and 1b). However, much of this variability was correlated in a region-specific fashion (Figure 2). In frontal areas, ΔPROG and ΔrCBF displayed a robust negative correlation (r=-0.82, p<0.01; figure 3a and 3b) whereas in the midbrain (substantia nigra or SN), a positive correlation (r=0.67,p<0.01; 3c and 3d) was observed. Interestingly ΔEST and arterial diameters were not correlated with ΔrCBF or ΔPROG, respectively (p>0.1).Discussion
To our knowledge, this is the first study to investigate how hormone fluctuations across the MC affect cerebral vascular function and structure. Preliminary results from this study show that PROG changes across the MC show a dose- and area-dependent relationship with CBF. In midbrain areas, progressive increases in PROG are associated with a proportional increase in CBF. In the frontal cortex, on the other hand, the same PROG increases were associated with a reduction in CBF. These findings indicate that PROG, as opposed to EST, is a potent modulator of CBF. Importantly, this was not driven by a change in the size of major feeding arteries, thus going against the notion that sex-hormones are primarily vasodilators. In addition, this effect is localized to the SN and frontal regions, suggesting that this circuit is particularly important in regulating hormone changes. Increased rCBF in the midbrain region may reflect changes in metabolic demand from local inhibitory interneurons controlling the SN dopaminergic neurons which project to the frontal cortex10,11. In fact, previous studies have shown that PROG can attenuate dopamine activity in the frontal cortex12,13. Taken together, our results demonstrate that PROG must be included as a variable when interpreting regional and sex differences in CBF.Conclusion
Findings from this study show that rCBF is dynamic across the MC. Further investigation is needed to clarify through which mechanisms PROG alters rCBF and how this affects other hemodynamic-based MRI measures, such as the BOLD signal. These results highlight the importance of considering not only sex but also MC phase in studies using hemodynamic based MRI sequences to control for the effect of the fluctuating PROG levels in the female brain on rCBF.Acknowledgements
Thank you, Éric Lavellée, Étienne Croteau, and Lyes Bachatene for your assistance with data collection.References
- Henriksen, O. M. et al. Sources of variability of resting cerebral blood flow in healthy subjects: A study using 133 Xe SPECT measurements. J. Cereb. Blood Flow Metab. 33, 787–792 (2013).
- Gur, R. E. & Gur, R. C. Gender differences in regional cerebral blood flow. Schizophr. Bull. 16, 247–254 (1990).
- Glezerman, M. Yes, there is a female and a male brain: Morphology versus functionality. Proc. Natl. Acad. Sci. 113, E1971–E1971 (2016).
- Beshay, V. E. & Carr, B. R. Clinical Reproductive Medicine and Surgery. 31–43 (2013). doi:10.1007/978-1-4614-6837-0
- Barbagallo, M. et al. Vascular effects of progesterone: Role of cellular calcium regulation. Hypertension 37, 142–147 (2001).
- Krause, D. N., Duckles, S. P. & Pelligrino, D. a. Influence of sex steroid hormones on cerebrovascular function. J. Appl. Physiol. 101, 1252–61 (2006).
- Chappell, M. A., Groves, A. R., Whitcher, B. & Woolrich, M. W. Variational Bayesian Inference for a Nonlinear Forward Model. IEEE Trans. Signal Process. 57, 223–236 (2009).
- Bizeau, A. et al. Stimulus-evoked changes in cerebral vessel diameter: A study in healthy humans. J. Cereb. Blood Flow Metab. 38, 528–539 (2018).
- Bernier, M., Cunnane, S. C. & Whittingstall, K. The morphology of the human cerebrovascular system. Hum. Brain Mapp. 1–14 (2018). doi:10.1002/hbm.24337
- 10. Zhang, Y., Michel-Herve Larcher, K., Misic, B. & Dagher, A. Anatomical and functional organization of the human substantia nigra and its connections. doi:10.7554/eLife.26653.001
- Gantz, S. C., Ford, C. P., Morikawa, H. & Williams, J. T. The Evolving Understanding of Dopamine Neurons in the Substantia Nigra and Ventral Tegmental Area. Annu. Rev. Physiol. 80, annurev-physiol-021317-121615 (2018).
- Thompson, T. L. & Moss, R. L. Modulation of mesolimbic dopaminergic activity over the rat estrous cycle. Neurosci. Lett. 229, 145–148 (1997).
- Grobin, A. C., Roth, R. H. & Deutch, A. Y. Regulation of the prefrontal cortical dopamine system by the neuroactive steroid 3a,21-dihidroxy-5a-pregnane-20-one. Brain Res. (1992).
- Allen Institute for Brain Science. Allen Brain Atlas. Webpage (2018). Available at: http://www.brain-map.org/. (Accessed: 22nd July 2018)
Figures
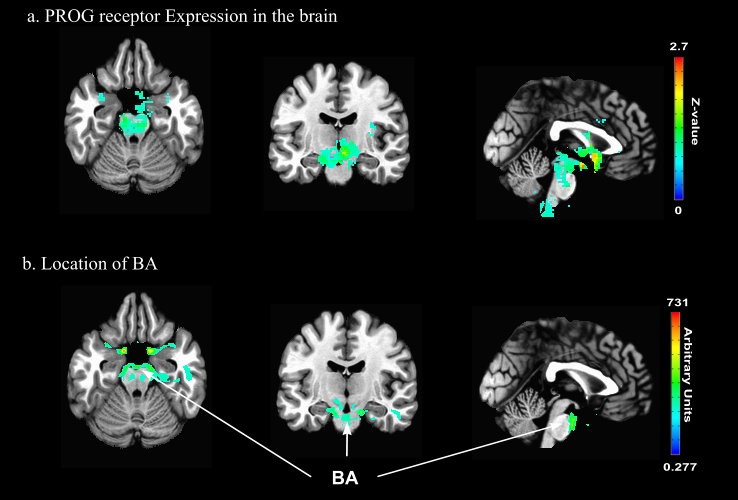
Figure 1. PROG receptors in the brain and location of major feeding vessels.
a. Location of PROG receptors in the human brain14 expressed as a z-score. Note that the highest expression is located in the midbrain and subcortical regions.
b. Location of the BA expressed in arbitrary units. Note the location of the BA with respects to the high concentration of PROG receptors.

Figure 2. Variability in PROG across the MC.
a. PROG levels in follicular and luteal phases of the MC, dashed lines denote women on oral contraception.
b. Variability in the change in serum ΔPROG, black diamonds denote women on oral contraception, black line denotes no change.

Figure 3. The effect of PROG in the FC and SN.
a. Correlation map of dose-dependent effects on CBF in the FC.
b. Scatter plot of average ΔCBF response in the FC and ΔPROG. Note that large increases in PROG lead to decreased CBF in the FC.
c. Correlation map of dose-dependent effects of PROG in the SN. Note the positive relationship between ΔCBF and ΔPROG.
d. Scatter plot of average ΔCBF response in the SN and ΔPROG. Women on oral contraceptives are denoted by black diamonds.

Figure 4. Proposed mechanism for PROG’s effect on CBF.
a. Dopaminergic pathway overlayed on the correlation map between ΔPROG and ΔCBF (no clustering or p-value threshold applied). Dopaminergic cell bodies originating in the SN have projections terminating throughout the frontal cortex where we observed decreases in CBF as PROG increased. Changes in cortical and midbrain CBF may reflect metabolic changes as GABAergic activity in the SN inhibits cortical dopamine release11.
b. Scatter plot of ΔFC CBF and ΔSN CBF. The more CBF increased in the SN when PROG increased, the more CBF in the FC decreased when PROG increased.