3233
Vulnerable Networks in the Aging Mouse Brain1Pratt School of Engineering, Duke University, Durham, NC, United States, 2Radiology, Duke Univ Medical Center, Durham, NC, United States, 3School of Informatics, Computing and Engineering, University of Indiana, Bloomington, IN, United States, 4Statistical Science, Duke University, Durham, NC, United States, 5Duke University Medical Center, Durham, NC, United States
Synopsis
Despite recent advances in aging research, the underlying mechanisms of selective brain vulnerability to aging remain to be elucidated. Mouse models may provide useful tools to dissect the mechanisms behind age and sex associated vulnerability of brain circuits. We used high resolution accelerated protocols and tensor network analyses to reveal structural network differences in aging C57BL/6 mice.
Introduction
A prelude to the study of network changes in mouse models of neurodegenerative disease needs to rely on understanding the network properties and our capabilities to study those in normative populations, as they undergo the aging process, as well as sex-based differences. As most models rely on the C57/BL6 background we sought to identify the age associated basis for brain circuit vulnerability using male and female C57BL/6 mice aged to 4 and 12 months, representative of young (~30 years old) to middle (~60 years old) human age.Methods
Animal groups of 10 mice at 4 months of age, and 20 mice at 12 months of age were evenly balanced for sex at each age. Brain specimens were fixed-perfused and enhanced with Gadolinium, before being imaged at 9.4T. To derive connectivity information, we used compressed sensing diffusion weighted protocols sensitized to 46 diffusion directions, interspersed with 5 non-diffusion weighted acquisitions. Images were acquired with TE=12 ms, TR=90 ms, b max ≈ 4000 s/mm2 , and we used a compression factor of 4, allowing for efficient sampling and reconstruction at 55 µm resolution, in a high performance computing environment1. The tracts connecting 332 atlas regions2 were used to build connectomes based on a constant solid angle (Q-Ball) method implemented in DIPY3. Tracts were visualized using MI-brain (imeka.ca). Network changes were estimated based on a recently proposed statistical method for dimensionality reduction4 using tensor network PCA. We hypothesized that age modulates network properties, and that we can identify vulnerable circuits in aging.Results
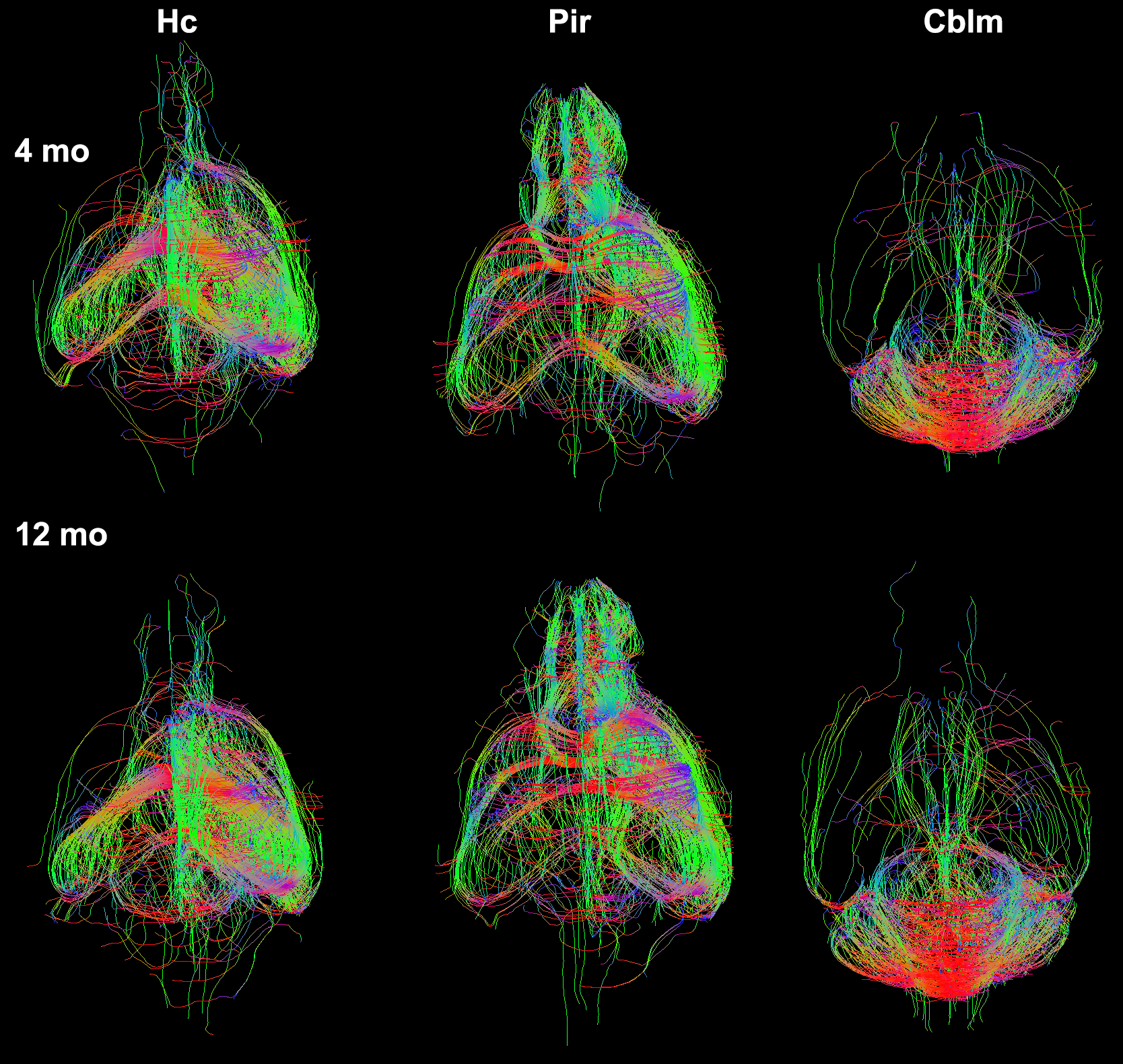
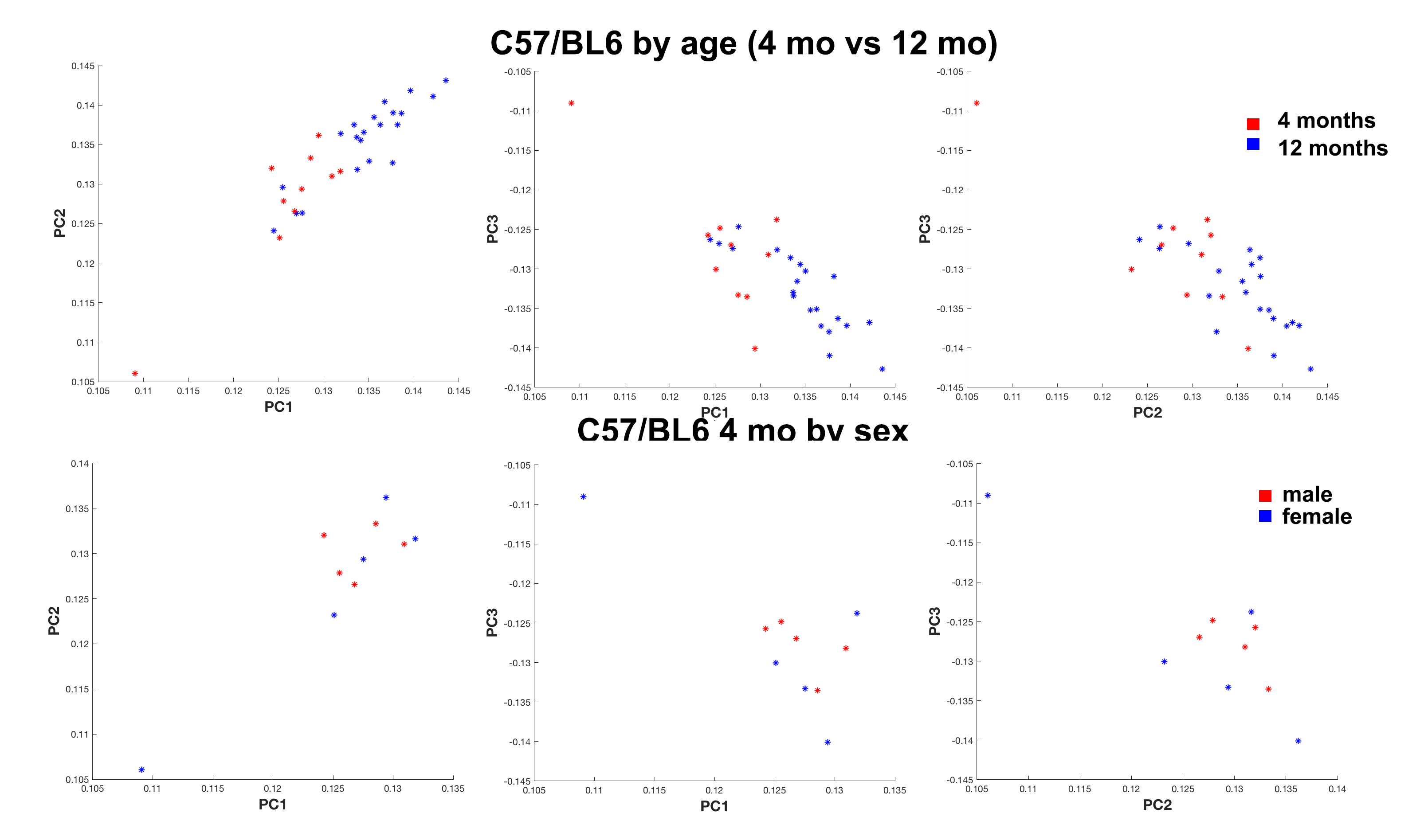
We have produced tractography and connectomes for the widely used C57/BL6 mouse strain at 4 and 12 months of age using efficient protocols based on compressed sensing, and we used a novel dimensionality reduction method called tensor network factorization, which relies on a generalization of principal component analysis4. Our results indicated that even though qualitative differences between representative animals of the two age groups were subtle (Figure 1), we could separate these groups based on a quantitative statistical analysis relying on the tensor network decomposition. However, sex differences were only apparent in the younger (4 months old) group (Figure 2).
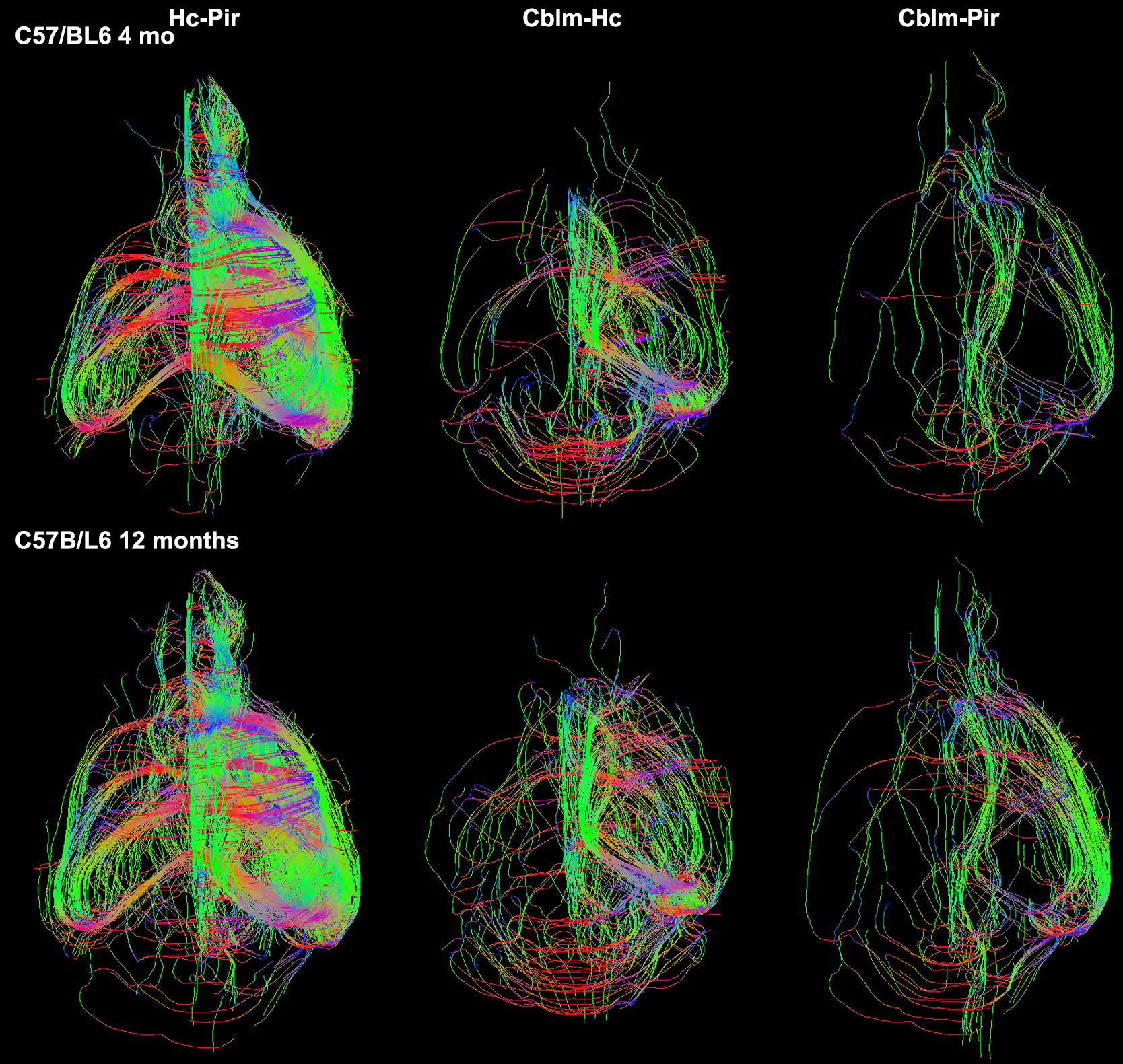
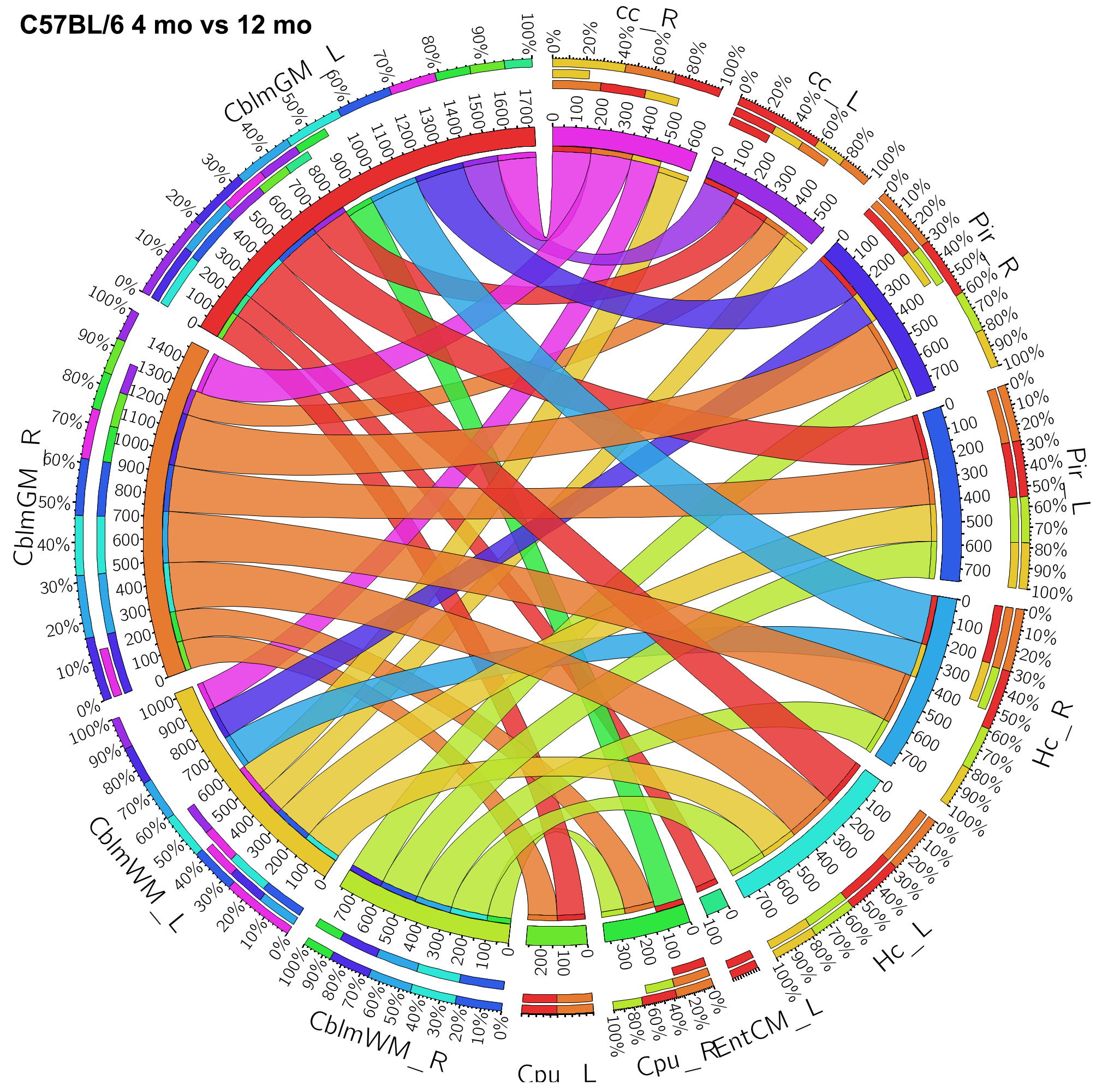
We identified the top ranked pairs of regions (out of 54780 connections) in terms of changes in connectivity with age. Our top 30 ranked results identified a role for the cerebellum, hippocampus, entorhinal cortex and piriform cortex, as well as for the cerebellar white matter and corpus callosum. Extending the list to the top 100 ranked pairs of connected regions helped identify an extended network comprised of 17 unique regions (13 gray matter regions, and 4 white matter regions) which contributed to distinguishing between the old and young groups. The gray matter regions included: accumbens, amygdala, caudomedial entorhinal cortex, cerebellar cortex, globus pallidus, hippocampus, hypothalamus, piriform cortex, preoptic telencephalon, septum, striatum, superior colliculus, and rest of thalamus. The white matter regions also included the inferior cerebellar peduncle and fimbria. The tracts reconstructed for the top connections are shown for representative specimens of the 4 months old and 12 months old mice groups in Figure 3. The group-wise network results are shown using chords diagrams in Figure 4. Our results suggest that regions commonly involved in age related neurodegeneration, as well as the cerebellum may play a role in age related vulnerability.
Discussion
Despite recent research advances, the underlying mechanisms of selective brain vulnerability to aging remain to be elucidated. We have provided connectivity data based on a compressed sensing acquisition in mouse models, to help elucidate the dynamics of age related vulnerability. While our data set is limited, expanding it to more animals and including both male and female mice may help us better understand the sex specific differences in relationship to vulnerability to aging.Acknowledgements
K01AG041211; R56AG051765; R56AG057895; P41EB015897References
1. Wang N, Anderson RJ, Badea A, Cofer G, Dibb R, Qi Y, Johnson GA. Whole mouse brain structural connectomics using magnetic resonance histology. Brain Struct Funct. 2018. Epub 2018/09/19. doi: 10.1007/s00429-018-1750-x. PubMed PMID: 30225830. 2. Anderson RJC, James J; Delpratt, Natalie A; Nouls, John C; Gu, Bin; McNamara, James O; Avants, Brian B; Johnson, G Allan; Badea, Alexandra. Small Animal Multivariate Brain Analysis (SAMBA): A High Throughput Pipeline with a Validation Framework. rXiv e-prints. 2017. 3. Garyfallidis E, Brett M, Amirbekian B, Rokem A, van der Walt S, Descoteaux M, Nimmo-Smith I. Dipy, a library for the analysis of diffusion MRI data. Front Neuroinform. 2014;8:8. PubMed PMID: 24600385. 4. Zhang Z, Allen GI, Hongtu Zhu H, Dunson D. Tensor network factorizations: Relationships between brain structural connectomes and traits. arXiv preprint arXiv:180602905. 2018.Figures