3208
Investigation of cerebral metabolite changes in the dorsolateral and ventrolateral prefrontal cortex of adult macaques with neonatal hippocampal lesions1Yerkes Imaging Center, Yerkes National Primate Research Center, Emory University, Atlanta, GA, United States, 2Department of Psychology and Yerkes National Primate Research Center, Emory University, Atlanta, GA, United States, 3Department of Bioengineering, University of California, Riverside, CA, United States, 4Division of Neuropharmacology and Neurologic Disease, Yerkes National Primate Research Center, Emory University, Atlanta, GA, United States
Synopsis
In the present study, in
vivo MR spectroscopy was employed to investigate the neuro-metabolites
changes of dorsolateral prefrontal cortex (DLPFC) and
ventrolateral prefrontal cortex (VLPFC) in adult macaques with neonatal
hippocampus lesion (Neo-H). Significant
metabolite changes were seen in the right DLPFC of
the Neo-H monkeys but not in the VLPFC, and significantly correlated
with the working memory scores. Also, lateralization of the cerebral metabolite
changes was observed. The results suggest that the neuro-metabolites changes in
DLPFC and VLPFC of adult monkeys resulted from early insult to the hippocampus
and the effect is mainly seen in the right hemisphere.
Introduction
Impaired memory is an important component of many developmental neuropsychiatric disorders in humans, such as schizophrenia (SZ), autism and stress disorders, that are often associated with medial temporal lobe dysfunction. Neonatal hippocampal (Neo-H) lesions in animals induce postpubertally manifested changes in prefrontal cortex (PFC) mimicking most aspects of SZ and has become the popular animal models in SZ researches [1-2]. Adult monkeys with Neo-H lesions have impaired working memory processes mediated by dorsolateral prefrontal cortex (DLPFC) but not by ventrolateral prefrontal cortex (VLPFC) [3]. Proton Magnetic Resonance Spectroscopy (1H-MRS) has been a valuable tool to examine the neurochemical changes in the brain of SZ [4]. Metabolite such as N-Acetyl Aspartate (NAA), Glutmate (Glu)/ Glutmin (Glx), and Choline levels in DLPFC and other brain areas have been used to assess the progression of the SZ disease [5]. However, few papers reported the metabolite changes in the animal models of SZ. In the present study, in vivo 1H-MRS were employed to investigate alterations of metabolite in DLPFC and VLPFC of adult monkeys with Neo-H lesions.Methods
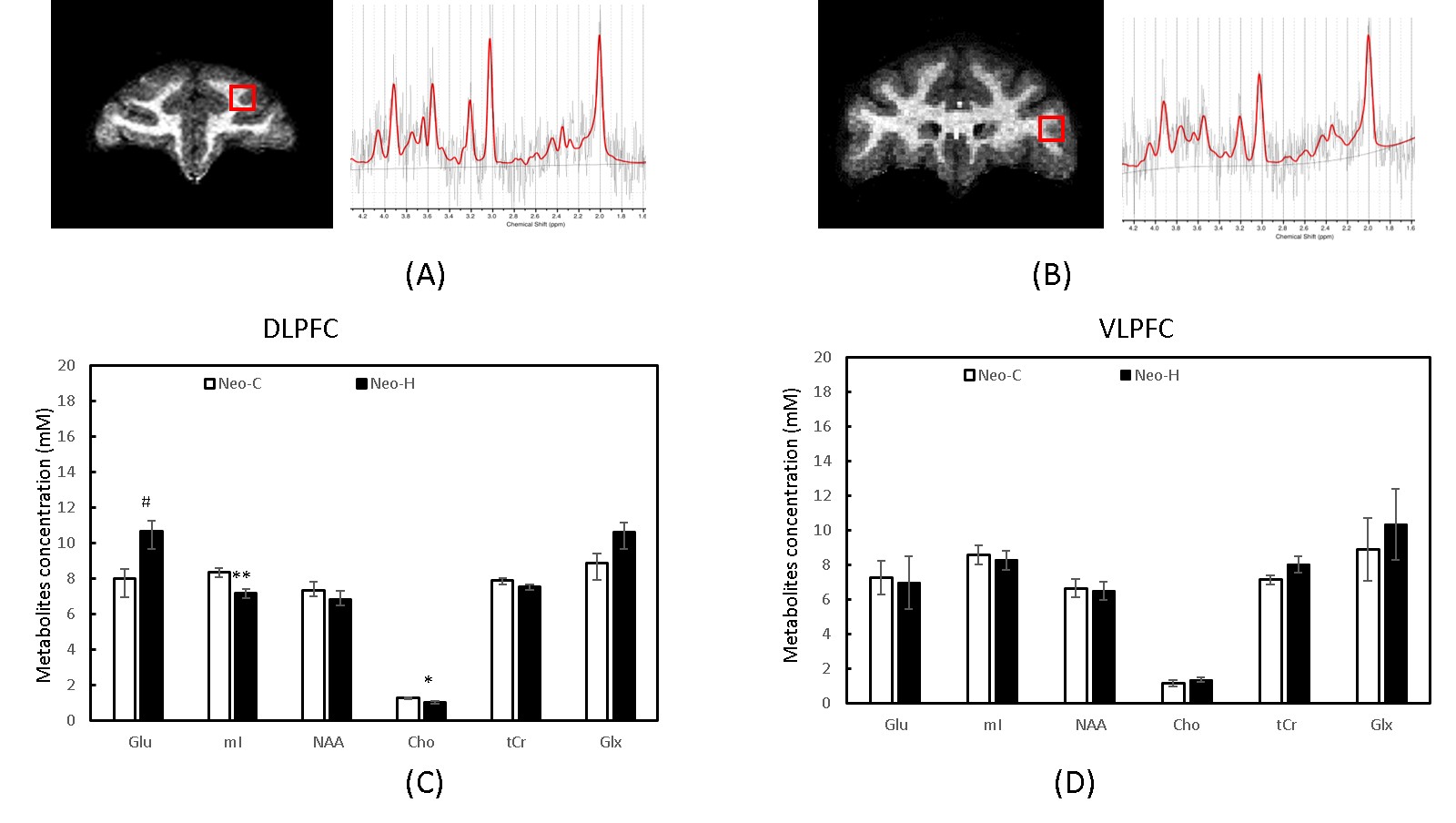
Adult rhesus macaques received either sham-operations (Neo-C, n=4) or neurotoxic lesions of the hippocampus (Neo-H, n=4) 10-15 days after birth. The MRS data were collected with a surface coil when the animals were 5-6 years old. Single voxel (5 × 5× 5 mm3) MRS was performed in left and right DLPFC and VLPFC (Fig 1. A and B) with a PRESS sequence with TR/TE =1500/30ms and CHESS water suppression. Concentration of metabolite, including NAA, creatine and phosphocreatine (tCr), total choline (tCho), myo-inositol (mI), and Glx, were derived from the spectra using the LC Model software (www.s-provencher.com) and the unsuppressed water peak was used as reference for absolute quantification. Working memory performance was previously assessed in all animals using the Object Self-Ordered (Obj-SO) and Session-Unique Delayed Nonmatching-to-Sample (SU-DNMS) tasks [3]. Independent sample t test was used to compare metabolite concentrations in selected ROIs between the two groups. Correlation between working memory scores (Obj-SO and SU-DNMS) and neuro-metabolite (NAA, mI, tCho, tCr, Glu and Glx) levels in the left and right DLPFC /VLPFC were investigated using Pearson correlation coefficient. P-values less than 0.05 were considered statistically significant in SPSS 24.0.Results
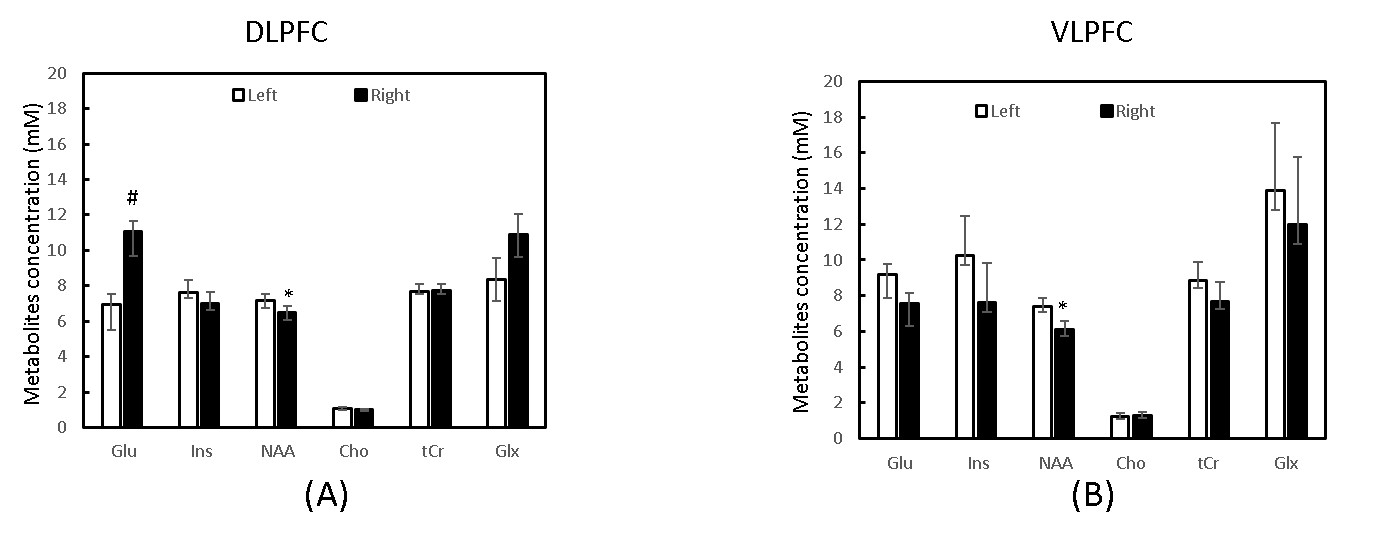
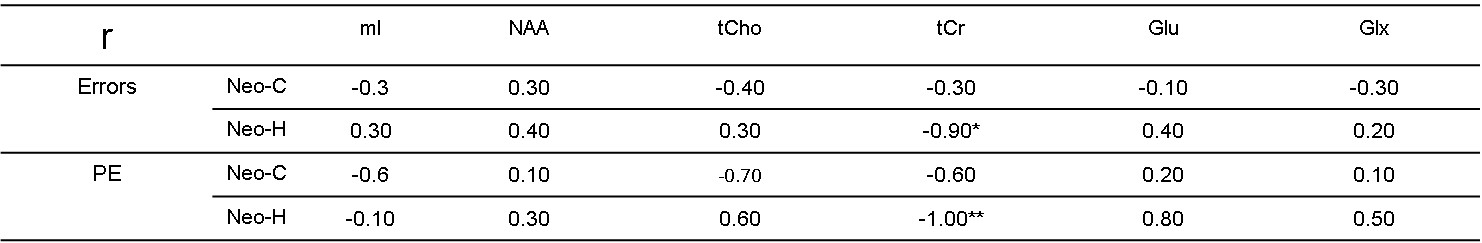
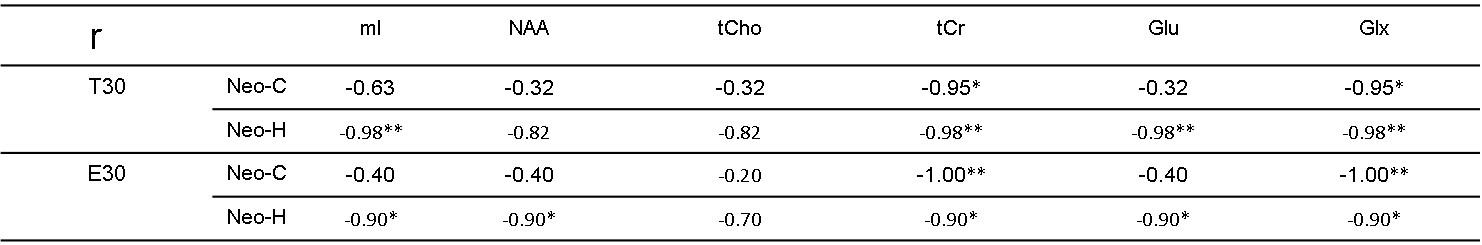
Significant metabolite changes were observed in the right DLPFC (Fig 1C). Compared to Neo-C, mI and tCho in right DLPFC were significantly decreased in monkeys with Neo-H, whereas Glx were dramatically increased (Fig.1C). No obvious metabolite changes were seen in VLPFC (Fig 1D). In addition, right lateralization was seen both in DLPFC and VLPFC of monkeys with Neo-H (Fig 2). There were significant correlations between metabolite changes of tCr in DLPFC and Obj-SO scores (Errors and total perseverative errors) (Table 1), and between metabolite changes (i.e. mI, NAA, tCr and Glx) in VLPFC and SU-DNMS scores (i.e. number of trials and errors at 30sec delays) (Table 2).Discussion and conclusion
We aimed to examine whether the neuro-metabolites would be affected in adult monkeys with Neo-H lesions and how changes in metabolites were correlated to cognitive performance. The present results showed decreased NAA in Neo-H monkeys (although not significant), which is consistent with the previous report that NAA levels decreased in the DLPFC of patients with SZ [6]. The significant correlation between tCr in right DLPFC and cognitive scores in Obj-SO indicated that the abnormal behavior severity is correlated with DLPFC dysfunction caused by Neo-H lesions in infancy. The data are consistent with prior findings of an association between tCr metabolism and cognitive function in SZ [7]. The difference between NAA changes in left and right DLPFC and VLPFC of Neo-H monkeys are also seen in prior study in AVH+ (auditory verbal hallucinations) patients of SZ [8].
In conclusion, the metabolite findings in the Neo-H monkey model are consistent with previous reports in SZ patients, so it is useful to investigate the mechanism of SZ disease.
Acknowledgements
This project was funded by NIH/NIMH
grant MH0588446 (JB), the National Center for Research Resources P51RR000165,
and the office of Research Infrastructure Programs/OD P51OD011132.
References
[1] A.M. Brady, The Neonatal Ventral Hippocampal Lesion (NVHL) Rodent Model of Schizophrenia, Curr Protoc Neurosci, 77 (2016) 9 55 51-59 55 17.
[2] X.S. Chen, C. Zhang, Y.F. Xu, M.D. Zhang, F.Y. Lou, C. Chen, J. Tang, Neonatal ventral hippocampal lesion as a valid model of schizophrenia: evidence from sensory gating study, Chin Med J (Engl), 125 (2012) 2752-2754.
[3] E. Heuer, J. Bachevalier, Neonatal hippocampal lesions in rhesus macaques alter the monitoring, but not maintenance, of information in working memory, Behav Neurosci, 125 (2011) 859-870.
[4] A. Schwerk, F.D. Alves, P.J. Pouwels, T. van Amelsvoort, Metabolic alterations associated with schizophrenia: a critical evaluation of proton magnetic resonance spectroscopy studies, J Neurochem, 128 (2014) 1-87.
[5] J.D. Port, N. Agarwal, MR spectroscopy in schizophrenia, J Magn Reson Imaging, 34 (2011) 1251-1261.
[6] N.V. Kraguljac, M. Reid, D. White, R. Jones, J. den Hollander, D. Lowman, A.C. Lahti, Neurometabolites in schizophrenia and bipolar disorder - a systematic review and meta-analysis, Psychiatry Res, 203 (2012) 111-125
[7] P.J. Allen, Creatine metabolism and psychiatric disorders: Does creatine supplementation have therapeutic value? Neurosci Biobehav Rev, 36 (2012) 1442-1462.
[8] M. Psomiades, M. Mondino, C. Fonteneau, R. Bation, F. Haesebaert, M.F. Suaud-Chagny, J. Brunelin, N-Acetyl-Aspartate in the dorsolateral prefrontal cortex in men with schizophrenia and auditory verbal hallucinations: A 1.5 T Magnetic Resonance Spectroscopy Study, Sci Rep, 8 (2018) 4133.
Figures