3196
In vivo longitudinal 1H MRS study of hippocampal, cereberal and striatal metabolic changes in the developing brain using an animal model of Chronic Hepatic Encephalopathy1LIFMET, CIBM, EPFL, Lausanne, Switzerland, 2CIBM, EPFL, Lausanne, Switzerland, 3Neurometabolic Unit, Service of Clinical Chemistry, University Hospital of Lausanne, Lausanne, Switzerland, 4Swiss Center for Liver Disease in Children, University Hospitals Geneva, Geneva, Switzerland
Synopsis
Chronic hepatic encephalopathy (CHE) is a severe neuropsychiatric disorder associated with chronic liver disease (CLD). For children, the impairment of neurocognitive functions as a consequence of CLD seems to be irreversible. We aimed to investigate longitudinally, using in-vivo 1H-MRS, differences in metabolic changes between hippocampus, cerebellum and striatum of a developing brain in a rat model of CHE. We showed the most pronounced changes in metabolites in cerebellum, suggesting its increased vulnerability. Further delineation of regional changes in the brain in response to CLD may help elucidate the molecular and regional origins of neuromotor and neurocognitive changes associated with CLD.
Introduction
Chronic hepatic encephalopathy (CHE) is a severe neuropsychiatric disorder associated with chronic liver disease (CLD) which can appear in both children and adults. For children with CLD the impairment of neurocognitive functions is described before liver transplantation and appears not to be reversible after the transplant indicating that the developing central nervous system could be more vulnerable to disease1-4. In addition, it has been reported, using an animal model of CLD-induced CHE that the hippocampus of a developing brain does show stronger metabolic changes compared to the adults5. Few previous studies in human patients with CHE at low B0 (1.5 T) evaluated brain regional differences in metabolic ratios of choline (Cho), glutamine and glutamate (Glx) and N-acetylaspartate (NAA) relative to creatine (Cr). Significant regional variations in these metabolite ratios were observed leading to a conclusion that information from different brain regions might be useful for better understanding of the mechanisms underlying the disease6. Thus, our hypothesis is that different brain regions will react differently to CLD-induced CHE.
To our knowledge, there are no published studies on developing brain during CLD assessing the potential brain regional differences. Therefore, the aim of this study is to investigate longitudinally, using in vivo 1H MRS, metabolic differences between hippocampus, cerebellum and striatum of a developing brain as key brain regions involved in a model of CHE7-9.
Methods
The bile duct ligation (BDL, accepted model of CHE) surgery was performed on 5 male Wistar rats at postnatal day 21 (P21). The maturation of rat brain at this age approximates a 9 month- old human10. BDL rats were always compared to sham operated animals at the same age (n=6) in order to: (1) consider the ongoing brain development and (2) take into account the well-known brain regional differences in healthy animals and therefore highlight only the brain regional difference due to disease.
Hippocampus (2x2.8x2mm3), cerebellum (2.5x2.5x2.5mm3) and striatum (2.5x2x2.5mm3) were scanned longitudinally using in vivo 1H MRS on 9.4 T system (Varian/Magnex Scientific). First scan, as well as blood tests, were performed at week2 after BDL and every two weeks up to week6 (scan2, 4, 6). MRS experiments were performed using the SPECIAL sequence (TE=2.8ms)11. First and second order shims were adjusted using FASTMAP12. Metabolite concentrations were calculated by LCModel using water as reference. Seventeen metabolites were reliable quantified, but only those showing a brain regional difference as a response to the disease are presented.
Results and Discussion
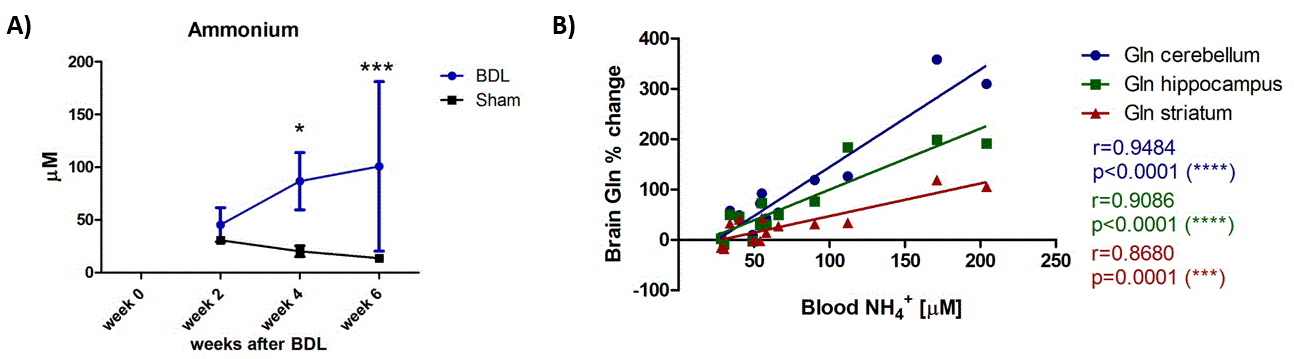
All the BDL operated rats showed increase in plasma bilirubin from week2 validating the presence of CLD. Increase in blood ammonia was also observed for all BDL rats when compared to the sham, reaching significance at week4 (Fig1 A). Increase in brain Gln concentration, as a response to NH4+ increase, was present in all three brain regions (week6: +173% in cerebellum, +112% in hippocampus, +68% in striatum) (Fig2). This increase was significant, compared to the shams, already at week4 in hippocampus and at week6 in cerebellum and striatum, although cerebellum showed the most pronounced increase in percentage at the end point of the disease (week6) (Fig2 B). Increase in blood ammonia correlated significantly with brain Gln for all brain regions, and the correlation was stronger in cerebellum (Fig1 B).
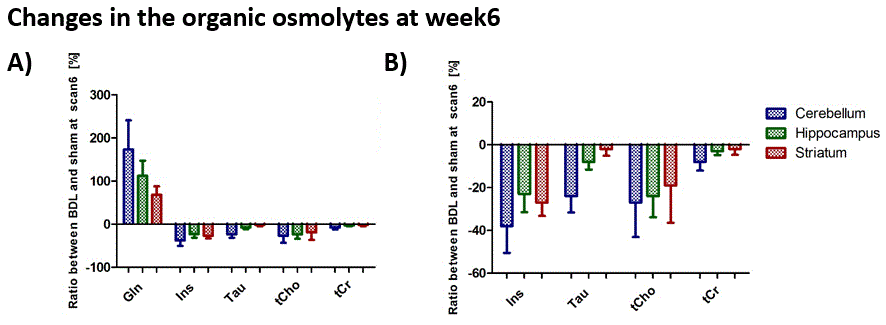
Main brain organic osmolytes decreased as a response to Gln increase (osmoregulation): taurine (Tau), total-creatine (tCr), total-choline (tCho) and myo-inositol (Ins). All of them displayed a tendency to stronger decrease in cerebellum at week6 (Fig3 A, B) except for tCho, probably due to stronger Gln increase in this region. Significant increase of lactate (Lac) concentration at the end of the disease (week6) was observed in all brain regions and again, it was stronger in cerebellum (+150% in cerebellum, +88% in hippocampus, +114% in striatum) indicating a possible energy metabolism perturbation (Fig4). This increase correlated significantly with the increase of brain Gln and blood NH4+ for all three mentioned brain regions. Neurotransmitter glutamate (Glu) and antioxidant ascorbate (Asc) also showed stronger decrease in cerebellum (data not shown).
Conclusion
This is the first study showing in vivo longitudinal overview of metabolic responses of the hippocampus, cerebellum and striatum to CLD in a developing brain. Overall, all three brain regions showed similar tendency of changes in metabolite concentrations. However, the changes in cerebellum seem to be the most pronounced suggesting increased vulnerability. Further delineation of regional changes in the brain in response to CLD may help elucidate the molecular and regional origins of neuromotor and neurocognitive changes associated with CLD.Acknowledgements
SNSF project no 310030_173222/1 and the CIBM (UNIL, UNIGE, HUG, CHUV, EPFL, the Leenaards and Jeantet Foundations) and CHUV and HUG.References
[1] Robertson C M T et al. Neurocognitive outcomes at kindergarten entry after liver transplantation at <3 yr. of age. Pediatric Transplantation 2013;17(7):621-630.
[2] Gilmour S M et al. School Outcomes in Children Registered in the Studies for Pediatric Liver Transplant (SPLIT) Consortium. Liver Transplantation 2010;16(9):1041-1048.
[3] Ng V et al. Development and validation of the pediatric liver transplantation quality of life: A disease-specific quality of life measure for pediatric liver transplant recipients. The Journal of Pediatrics 2014; 165(3):547-555.
[4] Sorensen L G et al. Longitudinal study of cognitive and academic outcomes after pediatric liver transplantation. The Journal of Pediatrics 2014;165(1):65-72.
[5] Rackayova et al. Proc. Intl. Soc. Mag. Reson. Med. 22 (2014) 0808.
[6] Taylor-Robinson, S D et al. Regional Variations in Cerebral Proton Spectroscopy in Patients with Chronic Hepatic Encephalopathy. Metabolic Brain Disease 1994;9(4):347-359.
[7] Arias N, Mendez M, Arias J L,Differential contribution of the hippocampus in two different demanding tasks at early stages of hepatic encephalopathy. Neuroscience 2015;284:1-10.
[8] Monfort P et al. Mechanisms of cognitive alterations in hyperammonemia and hepatic encephalopathy: Therapeutical implications. Neurochemistry International 2009;55(1-3):106-112.
[9] Weissenborn K et al. Attention, Memory and Cognitive Function in Hepatic Encephalopathy. Metab. Brain Disease 2005;20(4):358-367.
[10] Workman A et al. Modeling Transformations of Neurodevelopmental Sequences across Mammalian Species. Journal of Neuroscience 2013;33(17):7368-7383.
[11] Mlynárik V et al. Localized short- echo-time proton MR spectroscopy with full signal-intensity acquisition. Magnetic Resonance in Medicine 2006;56(5):965-970.
[12] Gruetter R, Tkác I, Field mapping without reference scan using asymmetric echo-planar techniques. Magnetic Resonance in Medicine 2000;43(2):319-323.
Figures