3192
Diffusion tensor imaging and resting-state functional MRI reveal altered brain network hubs on a depression knockout mouse model1Institute of Biomedical Engineering and Nanomedicine, National Health Research Institutes, Miaoli, Taiwan, 2Department of Biomedical Engineering and Environmental Sciences, National Tsing Hua University, Hsinchu, Taiwan, 3Graduate Institute of Pathology, National Taiwan University College of Medicine, Taipei, Taiwan, 4Department of Physics, National Taiwan University, Taipei, Taiwan, 5Department of Pathology, National Taiwan University Hospital, Taipei, Taiwan, 6Institute of Medical Device and Imaging, National Taiwan University College of Medicine, Taipei, Taiwan
Synopsis
In this study, diffusion tensor imaging (DTI) and resting-state functional MRI were employed on a depression knock-out mouse model, which shows behaviors of anxiety and depression. The brain network hubs were investigated by region-of-interest (ROI) and connectivity analyses. Our results showed altered resting-state connectivity in prefrontal and hippocampal areas. Also, altered DTI indices were also found in thalamus and hippocampus. These findings are consistent with previous human studies and suggest the brain neuroimaging could be potentially useful to reveal the brain network hubs affected by depression on the proposed mouse model.
Introduction
Depression is one of the most common psychiatric disorders worldwide, which could cause severe health conditions to the people affected. Recently, a BLM-s gene knockout mouse model has been developed with behavioral phenotype of anxiety and depression. To further understand the underlying mechanism of the brain network affected by depression, in this study, we utilized diffusion tensor imaging (DTI) and resting-state functional MRI (rs-fMRI) on this knockout mouse model and examined the intrinsic connectivity across different network hubs.Methods
All experiments were approved by the animal ethics review committee. A total of 12 wild-type (WT) mice and 10 BLM-s gene knockout mice were scanned on a 7T animal MRI (Bruker Biospec, Germany). Anesthesia was carried out by dexmedetomidine (0.5 mg/kg body weight) via i.p. injection. Respiratory rate was monitored and body temperature was kept with warm water circulation. To minimize head motion, 1% isoflurane anesthesia was introduced during DTI scan. The rs-fMRI data was acquired by using gradient-echo EPI with the following parameters: FOV=20×20 mm2, matrix size=80×80, TR/TE=2000/15 ms, 250 time points. With the same geometrical setting as rs-fMRI, the DTI data consisting of 5 b0 images and 30 diffusion-weighted images was acquired with the following parameters: matrix size=128×128, TR/TE=3750/31.2 ms, average=4 and b=1000 s/mm2. Upon the completion of MR experiments, the animals with antidote administration were placed in a cage with warm light till recover. The pre-processing steps including motion correction, image coregistration, and spatial smoothing (0.5 mm Gaussian) were carried out using SPM12 (UCL, UK). The detrending and bandpass filtering (0.01 Hz – 0.1 Hz) were applied by using REST toolkit (http://restfmri.net/forum/index.php). The resting-state networks (RSNs) were determined through seed-based analysis by in-house Matlab script, performing pixel-wise calculation of Pearson’s correlation coefficient (CC). The CC values among 11 representative network hubs 1 were also assessed. For analyzing DTI, DSI studio (http://dsi-studio.labsolver.org/) was used to calculate the diffusion quantitative indices for following statistical analysis.Results
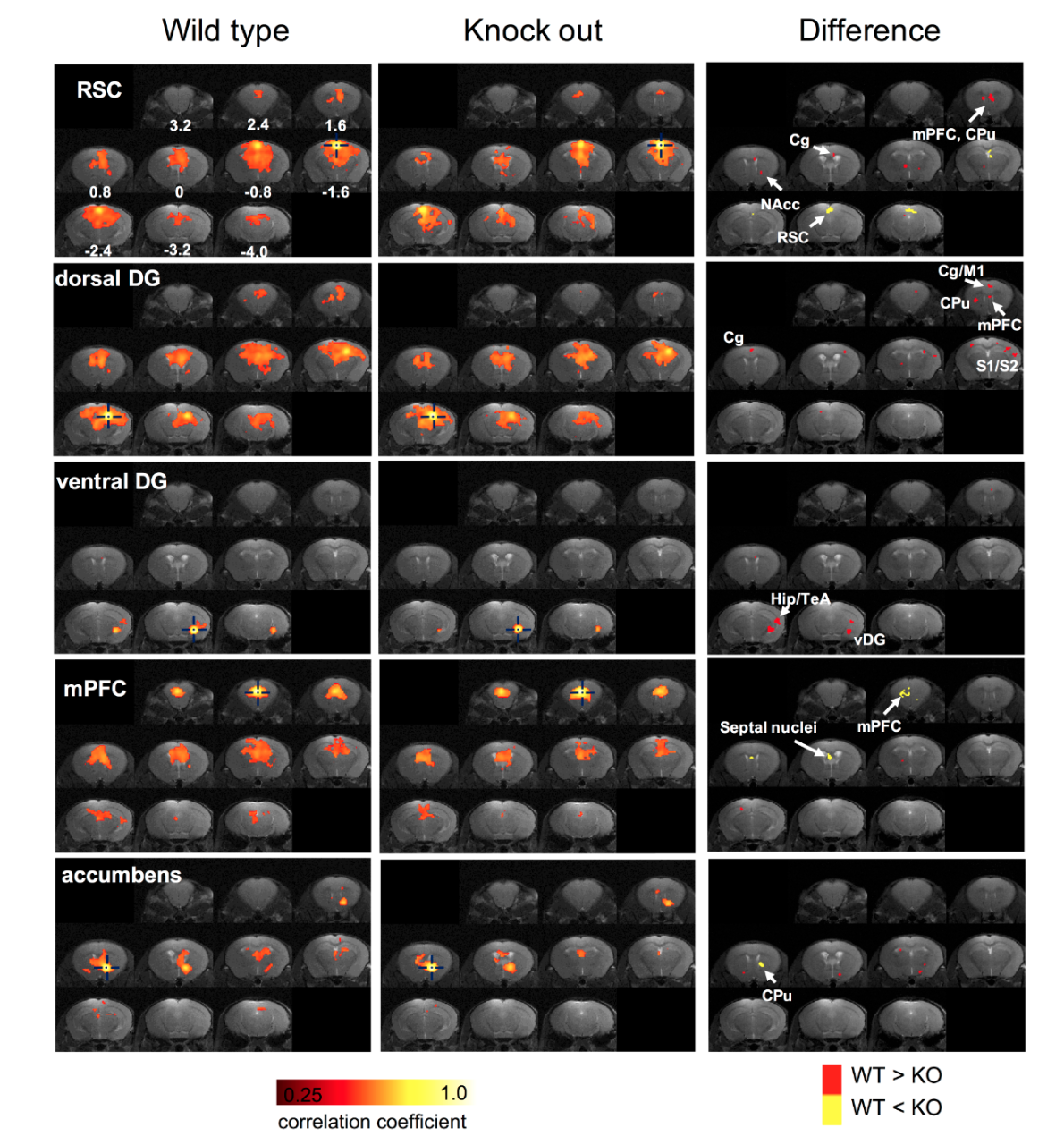
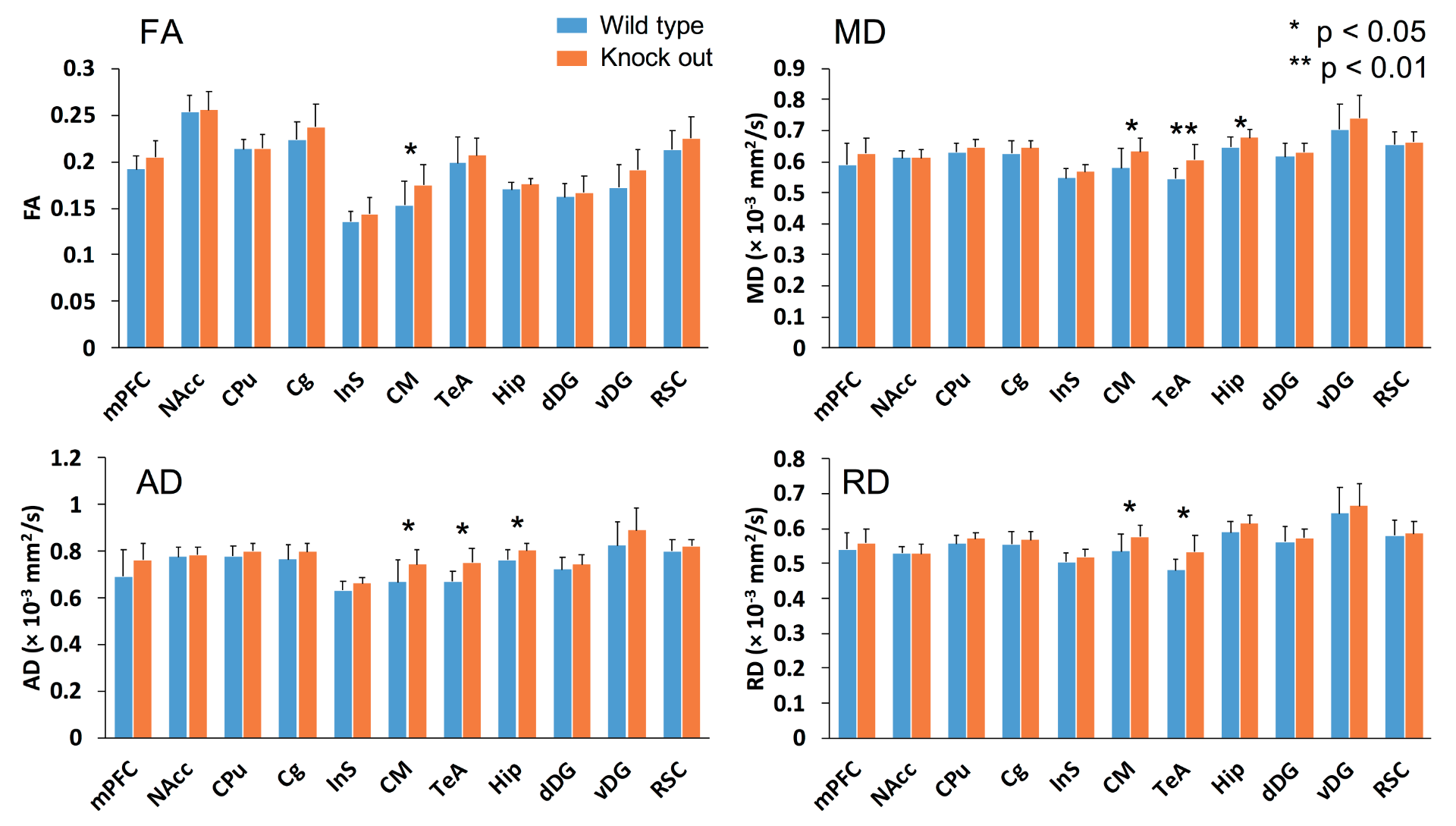
Figure 1 shows the comparison of five RSNs between wide-type and knockout mice. The significant differences were found in medial prefrontal cortex (mPFC), cingulate cortex (Cg), hippocampal area, caudate putamen (CPu), septal nuclei, and retrosplenial cortex (RSC). Figure 2 shows the connectivity matrices among the 11 region-of-interest (ROIs) for both groups. As depicted in the difference map, the mPFC of WT mice presents higher connectivity in hippocampus and ventral dentate gyrus (DG). Interestingly, several significant differences were found concerning to ventral DG. A comparison of DTI quantitative indices was shown in figure 3, revealing signifcant differences in central medial nuclei (CM), temporal association cortex (TeA) and hippocampus.Discussions
In a previous human study, increased connectivity in dorsomedial prefrontal cortex and precuneous of DMN has been observed in depression subjects 2. Consistently, our results show that the DMN-like network of knockout mice possessed an increased connectivity within RSC region. Besides, reduced connectivity between mPFC and hippocampal areas in knockout mice may imply the dysfunction on this circuit. Although previous rs-fMRI studies on patients regarding to mPFC-hippocampus connectivity were not consistent 3,4, our findings show that altered mPFC-hippocampus connectivity could be observed in the mouse model. As a central hub of thalamic network, mapping the functional connectivity related to CM is beneficial to understand the role of thalamic network in this model. Although no significant difference was found related to the rs-fMRI connectivity of CM nucleus, DTI shows substantial differences of FA and diffusivity between wide-type and knockout groups. Consistently, thalamic abnormalities have been previously found associated with depression through rs-fMRI 5-7. Therefore, the roles of CM nucleus as well as thalamic area in this knockout mouse model have to be investigated in further studies. Another interesting finding has been found in hippocampal region. Our rs-fMRI analysis shows differences of connectivity regarding to ventral DG, indicating the functional impairment of ventral DG or ventral hippocampal area. The DTI analysis also shows that the microstructural properties in hippocampal region were altered in the knockout mouse group. Functionally, it has been suggested that ventral part of the hippocampus is in charge of emotional and affective regulation 8,9, which may be associated with depression. Thus, the possible origin of the hypo-activity in ventral DG may play an important role in understanding the etiology of depression as well as developing therapeutic approaches.Conclusion
In summary, we utilized DTI and rs-fMRI techniques on a depression knockout mouse model and examined the altered connectivity as well as diffusion characteristics. Our results show that the abnormalities can be observed in particularly mPFC and ventral hippocampal regions, demonstrating the potential of the use of neuroimaging on this knockout mouse model. Further histological studies are needed to verify the neuroimaging results and solidifying the possible origins of alterations.Acknowledgements
We thank for the funding supports from National Health Research Institutes (BN-107-PP-06), Taiwan Ministry of Science and Technology (107-2321-B-400-005 and 107-2221-E-400-001), and the Taiwan Central Government S & T grant (107-1901-01-19-02).References
1. Liska A, Galbusera A, Schwarz AJ, Gozzi A. Functional connectivity hubs of the mouse brain. Neuroimage 2015;115:281-291.
2. Sheline YI, Price JL, Yan ZZ, Mintun MA. Resting-state functional MRI in depression unmasks increased connectivity between networks via the dorsal nexus. P Natl Acad Sci USA 2010;107(24):11020-11025.
3. Goveas J, Xie CM, Wu ZL, Ward BD, Li WJ, Franczak MB, Jones JL, Antuono PG, Yang Z, Li SJ. Neural correlates of the interactive relationship between memory deficits and depressive symptoms in nondemented elderly: Resting fMRI study. Behav Brain Res 2011;219(2):205-212.
4. Geng H, Wu F, Kong L, Tang Y, Zhou Q, Chang M, Zhou Y, Jiang X, Li S, Wang F. Disrupted Structural and Functional Connectivity in Prefrontal-Hippocampus Circuitry in First-Episode Medication-Naive Adolescent Depression. PLoS One 2016;11(2):e0148345.
5. Yamamura T, Okamoto Y, Okada G, Takaishi Y, Takamura M, Mantani A, Kurata A, Otagaki Y, Yamashita H, Yamawaki S. Association of thalamic hyperactivity with treatment-resistant depression and poor response in early treatment for major depression: a resting-state fMRI study using fractional amplitude of low-frequency fluctuations. Transl Psychiat 2016;6.
6. Greicius MD, Flores BH, Menon V, Glover GH, Solvason HB, Kenna H, Reiss AL, Schatzberg AF. Resting-state functional connectivity in major depression: abnormally increased contributions from subgenual cingulate cortex and thalamus. Biol Psychiatry 2007;62(5):429-437.
7. Anand A, Li Y, Wang Y, Wu J, Gao S, Bukhari L, Mathews VP, Kalnin A, Lowe MJ. Activity and connectivity of brain mood regulating circuit in depression: a functional magnetic resonance study. Biol Psychiatry 2005;57(10):1079-1088.
8. Fanselow MS, Dong HW. Are the dorsal and ventral hippocampus functionally distinct structures? Neuron 2010;65(1):7-19.
9. Moser MB, Moser EI. Functional differentiation in the hippocampus. Hippocampus 1998;8(6):608-619.
Figures