3044
Longitudinal characterization of white matter degeneration in a rat model of brain glucose hypometabolism and sporadic Alzheimer's disease1Faculdade de Ciências da Universidade de Lisboa, Lisbon, Portugal, 2Centre d'Imagerie Biomédicale, Ecole Polytechnique Fédérale de Lausanne, Lausanne, Switzerland
Synopsis
Impaired brain glucose consumption is a possible trigger of Alzheimer’s disease (AD). Animal models can help characterize each contributor to the cascade independently. Here we use the intracerebroventricular-streptozotocin rat model of AD in a first-time longitudinal study of white matter degeneration using diffusion MRI. Diffusion and kurtosis tensor metrics reveal alterations in the cingulum, fimbria and fornix. The two-compartment WMTI-Watson biophysical model further characterizes the cingulum damage as axonal injury and loss - consistent with previous histopathological studies. White matter degeneration induced by brain glucose metabolism disruption can bring further insight into the role of this mechanism in AD.
Introduction
The mechanism that triggers Alzheimer’s disease (AD) is not well-established, with amyloid plaques, neurofibrillary tangles of tau, microgliosis and glucose hypometabolism all likely involved in the cascade. One main advantage of animal models is the possibility to tease out the impact of each of these insults on the neurodegeneration.
Following an intracerebroventricular (icv) injection of streptozotocin (STZ), rats and monkeys develop impaired brain glucose metabolism1 (i.e. “diabetes of the brain”2). Numerous studies have reported AD-like features in icv-STZ animals3,4,5, but this model has never been characterized in terms of MRI-derived biomarkers beyond structural brain atrophy4.
White matter (WM) degeneration has been proposed as a promising biomarker for AD that well precedes cortical atrophy and correlates strongly with disease severity6. Here, we performed a longitudinal study of WM degeneration in icv-STZ rats using diffusion MRI. White matter microstructure was characterized in terms of diffusion and kurtosis tensor metrics. The WMTI-Watson two-compartment model7 was further estimated to provide a more specific assessment of WM damage induced by glucose hypometabolism.
Methods
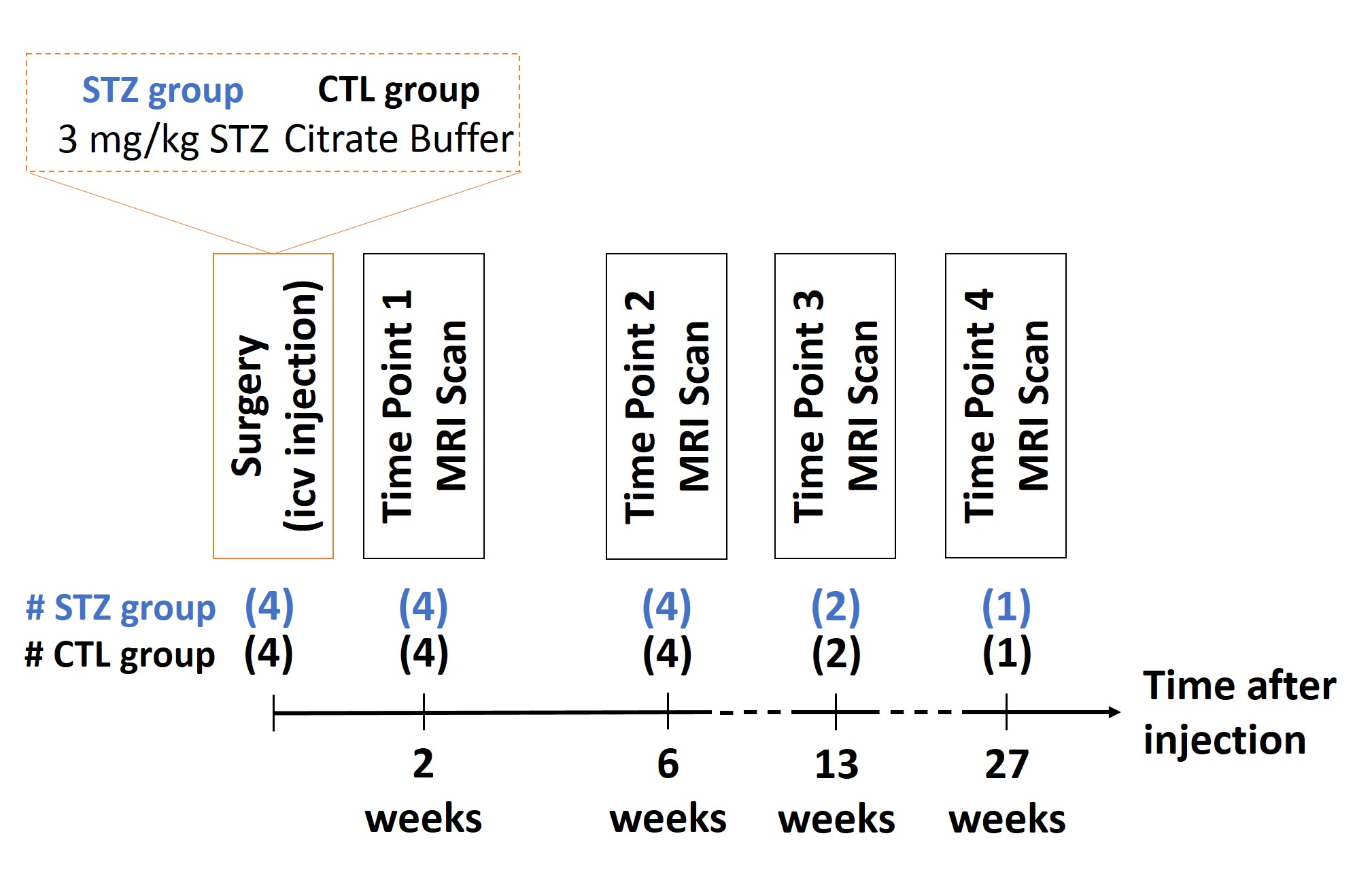
All experiments were approved by the local Service for Veterinary Affairs. Male Wistar rats (N=8) (236±11g) underwent a bilateral icv-injection of either streptozotocin (3mg/kg, STZ group, N=4) or buffer (control group, N=4). Rats were scanned at four timepoints following surgery (Fig.1), on a 14T Varian system. Diffusion data were acquired using a semi-adiabatic SE-EPI PGSE sequence as follows: 4 (b=0), 12 (b=0.8ms/µm2), 16 (b=1.3ms/µm2) and 30 (b=2ms/µm2) directions; TE/TR=48/2500ms, 9 coronal 1mm slices, δ/Δ=4/27ms, FOV=23x17mm2, matrix=128x64, 4 shots.
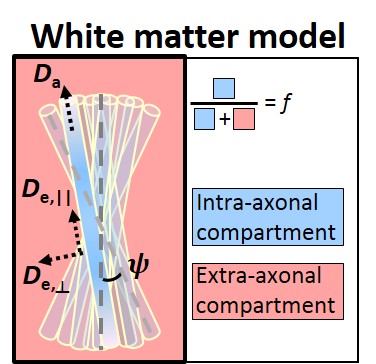
Images were denoised8 and EDDY-corrected9. Diffusion and kurtosis tensors were calculated for each voxel10, producing parametric maps of fractional anisotropy (FA), mean, axial and radial diffusivity (MD, AxD and RD) and mean, axial and radial kurtosis (MK, AK and RK). Additionally, five metrics were derived from the WMTI-Watson model (Fig.2) for WM voxels. Since the model allows for two mathematical solutions11, the Da>De,|| solution was retained based on recent evidence12.
Considering previous findings1,13, corpus callosum (CC), cingulum, fornix and fimbria were chosen as WM ROIs and automatically segmented using atlas-based registration. Mean diffusion metrics were calculated in each ROI for each dataset. To minimize partial volume effects with CSF, particularly marked in the STZ group due to structural atrophy and ventricle enlargement, voxels with MD>2.5μm2/ms were excluded. For the WMTI-Watson model, only voxels with estimates within biological ranges were retained.
Control (CTL) and STZ groups were compared using two-sided t-tests at each timepoint. Within-group longitudinal changes were assessed using one-way ANOVA and Tukey-Cramer correction for multiple comparisons. Because of the small cohort, t-tests were only performed at 2 and 6-week timepoints and ANOVA excluded the last timepoint.
Results
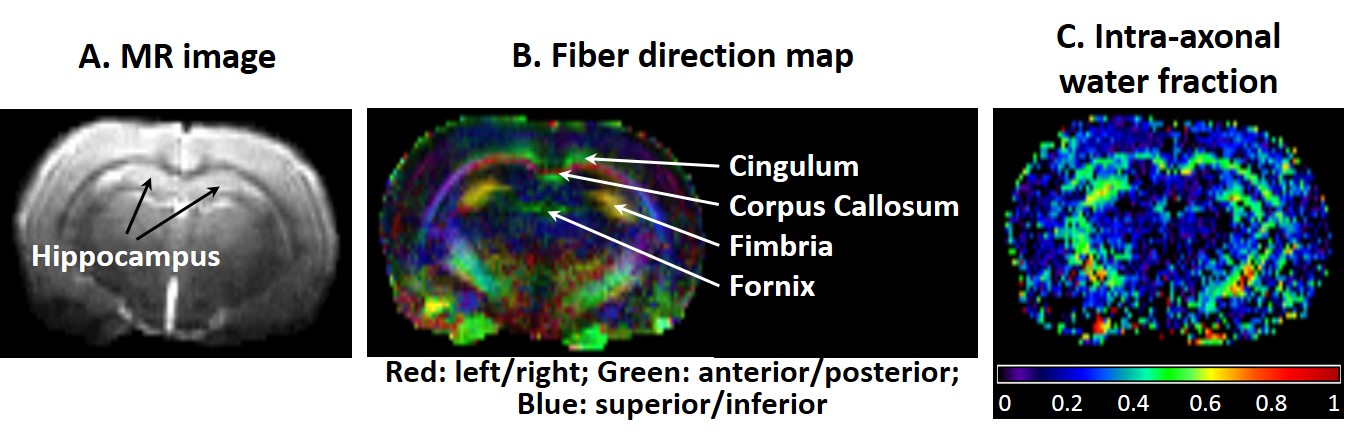
Examples of a b=0 image, color-coded FA and intra-axonal water fraction maps are shown in Fig.3.
Significant
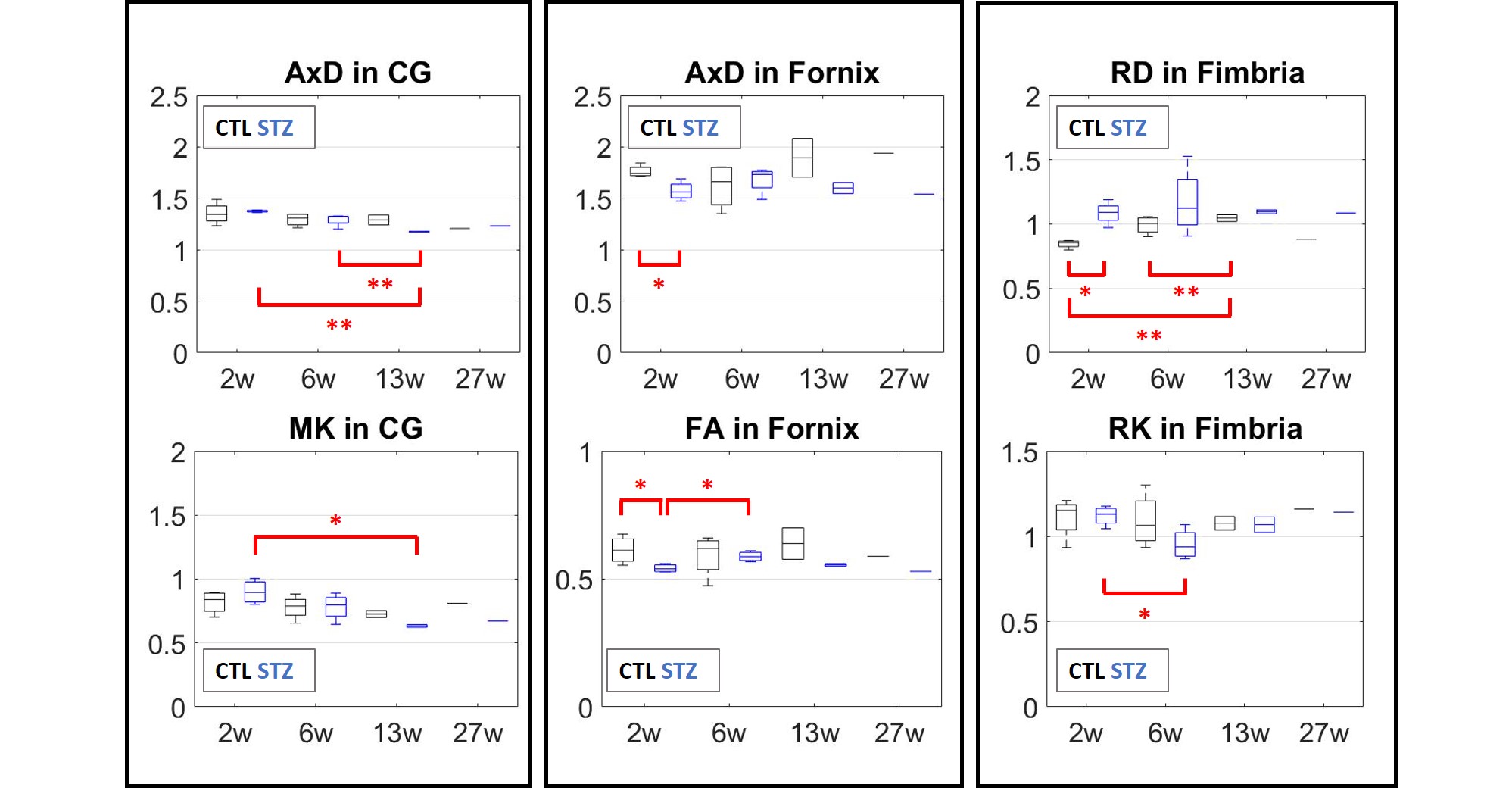
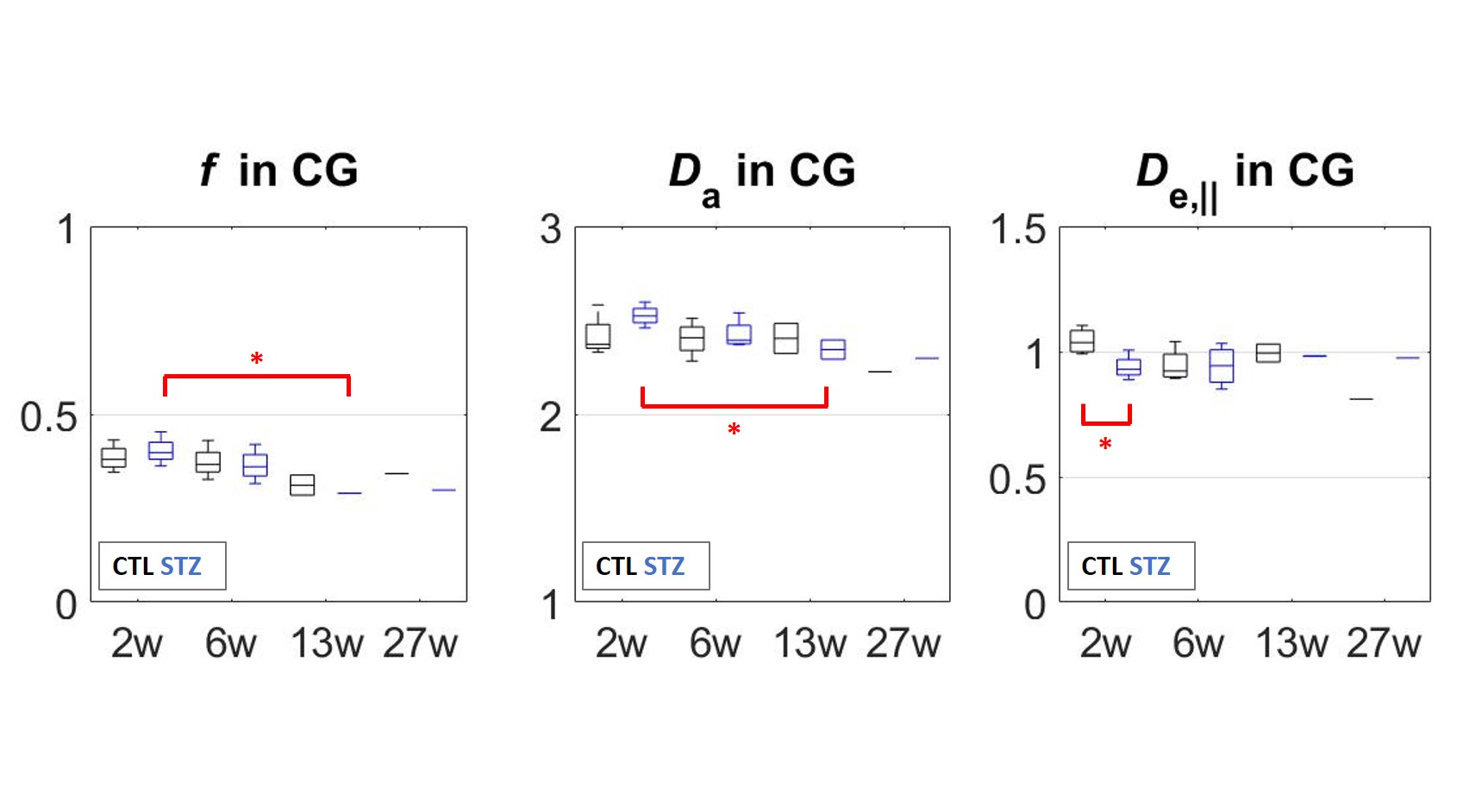
changes in DTI and DKI metrics are collected in Fig.4. In terms of WMTI-Watson metrics (Fig.5), the cingulum was the only ROI to display significant differences.
No significant differences were found in the CC, but a larger cohort might be necessary. The study is on-going, thus more data is being acquired.
Discussion and Conclusions
WM microstructure alterations were consistent with previous studies of the icv-STZ rat model using histopathology. Changes were most pronounced in the cingulum and suggested axonal damage and loss as early as six weeks. Since the cingulum connects parahippocampal gyrus to posterior cingulate and parietal cortices, our results are consistent with reported structural thinning of the parietal cortex after one month and intra-cellular accumulation of amyloid in the parietal cortex and hippocampus after 3 months3. Our measured changes in the fornix and fimbria are also consistent with reported atrophy of hippocampus and fornix, and concomitant myelin alterations and inflammation in the fornix at 6 weeks5. Finally, we report non-monotonic changes in the fimbria and fornix of STZ group, with partial recovery around 6 weeks, consistent with previously-established trends for structural thickness and memory performance3.
In the STZ group, a larger number of voxels failed to produce an acceptable solution under the Da>De,|| assumption enforced by the model. It is possible that the other solution (Da<De,||) better suits the pathological group. Future work will focus on finding the best fitting solution, and acquiring a larger dataset. We will also characterize gray matter degeneration using both diffusion and functional MRI and correlate it to WM degeneration.
Finally, the icv-STZ model displays WM changes in tracts also affected by AD, while the degeneration is induced primarily by impaired brain glucose metabolism. It therefore constitutes an excellent model of sporadic AD and should allow to further explore the hypothesis of AD being “type-3 diabetes”.
Acknowledgements
The authors thank Mario Lepore and Stefan Mitrea for assistance with animal setup and monitoring. This work was supported by the Centre d'Imagerie Bio-Médicale (CIBM) of the University of Lausanne (UNIL), the Swiss Federal Institute of Technology Lausanne (EPFL), the University of Geneva (UniGe), the Centre Hospitalier Universitaire Vaudois (CHUV) and the Hôpitaux Universitaires de Genève (HUG), the Swiss-European Mobility Plan (Movetia) and the Leenaards and the Jeantet Foundations.References
1. Hoyer, S., Nitsch, R., Oesterreich, K., 1991. Predominant abnormality in cerebral glucose utilization in late-onset dementia of the Alzheimer type: a cross-sectional comparison against advanced late-onset and incipient early-onset cases. J Neural Transm Park Dis Dement Sect 3, 1-14.
2. Steen, E., Terry, B.M., Rivera, E.J., Cannon, J.L., Neely, T.R., Tavares, R., Xu, X.J., Wands, J.R., de la Monte, S.M., 2005. Impaired insulin and insulin-like growth factor expression and signaling mechanisms in Alzheimer's disease--is this type 3 diabetes? J Alzheimers Dis 7, 63-80.
3. Knezovic, A., Osmanovic-Barilar, J., Curlin, M., Hof, P.R., Simic, G., Riederer, P., Salkovic-Petrisic, M., 2015. Staging of cognitive deficits and neuropathological and ultrastructural changes in streptozotocin-induced rat model of Alzheimer's disease. J Neural Transm (Vienna) 122, 577-592.
4. Kraska, A., Santin, M.D., Dorieux, O., Joseph-Mathurin, N., Bourrin, E., Petit, F., Jan, C., Chaigneau, M., Hantraye, P., Lestage, P., Dhenain, M., 2012. In vivo cross-sectional characterization of cerebral alterations induced by intracerebroventricular administration of streptozotocin. PLoS One 7, e46196.
5. Shoham, S., Bejar, C., Kovalev, E., Weinstock, M., 2003. Intracerebroventricular injection of streptozotocin causes neurotoxicity to myelin that contributes to spatial memory deficits in rats. Exp Neurol 184, 1043-1052.
6. Chang, Y.L., Chen, T.F., Shih, Y.C., Chiu, M.J., Yan, S.H., Tseng, W.Y., 2015. Regional cingulum disruption, not gray matter atrophy, detects cognitive changes in amnestic mild cognitive impairment subtypes. J Alzheimers Dis 44, 125-138.
7. Jespersen, S.N., Olesen, J.L., Hansen, B., Shemesh, N., 2018. Diffusion time dependence of microstructual parameters in fixed spinal cord. Neuroimage 182, 329-342.
8. Veraart, J.; Fieremans, E. & Novikov, D.S. Diffusion MRI noise mapping using random matrix theory Magn. Res. Med., 2016, early view, doi:0.1002/mrm.26059
9. Jesper L. R. Andersson and Stamatios N. Sotiropoulos. An integrated approach to correction for off-resonance effects and subject movement in diffusion MR imaging. NeuroImage, 125:1063-1078, 2016.
10. Veraart J, Sijbers J, Sunaert S, Leemans A, Jeurissen B. Weighted linear least squares estimation of diffusion MRI parameters: strengths, limitations, and pitfalls. Neuroimage 2013; 81: 335–346.
11. Jelescu, I.O., Veraart, J., Fieremans, E., Novikov, D.S., 2016b. Degeneracy in model parameter estimation for multi-compartmental diffusion in neuronal tissue. NMR Biomed 29, 33-47.
12. Kunz, N., da Silva, A.R., Jelescu, I.O., 2018. Intra- and extra-axonal axial diffusivities in the white matter: Which one is faster? Neuroimage 181, 314-322.
13. Fieremans, E., Benitez, A., Jensen, J.H., Falangola, M.F., Tabesh, A., Deardorff, R.L., Spampinato, M.V., Babb, J.S., Novikov, D.S., Ferris, S.H., Helpern, J.A., 2013. Novel white matter tract integrity metrics sensitive to Alzheimer disease progression. AJNR Am J Neuroradiol 34, 2105-2112.
Figures