2594
Altered cerebellar functional connectivity in Parkinson’s disease1Center for Advanced Neuroimaging, University of California Riverside, Riverside, CA, United States, 2Department of Neurology, Emory University, Atlanta, GA, United States, 3Department of Bioengineering, University of California Riverside, Riverside, CA, United States
Synopsis
Parkinson’s disease is a progressive, neurodegenerative disorder characterized by asymmetrical onset of motor symptoms such as bradykinesia, rigidity, and tremor. Mounting evidence suggests that the cerebellum plays a major role in the pathophysiology of PD. Prior imaging studies have found altered cerebellar activation during motor execution and motor learning, suggesting that altered activation in the cerebellum may reflect Parkinsonian-related impairment. Here, we use resting-state function MRI (fMRI) to ascertain connectivity changes in the cerebellum from Parkinson's disease found reduced connectivity in lobule V of the cerebellum as well as reduced connectivity between dentate nucleus and the cerebellar cortex.
Introduction
Parkinson’s disease (PD) is a progressive, neurodegenerative disorder characterized by asymmetrical onset of motor symptoms such as bradykinesia, rigidity, and tremor. The principal pathology in PD is the loss of melanized dopamine neurons in the substantia nigra pars compacta, and much of the work examining PD pathology has focused on ascertaining changes in substantia nigra and basal ganglia1. However, mounting evidence suggests that the cerebellum plays a major role in the pathophysiology of PD. For example, interruptions in the cerebello-thalamo-cortical circuit may be involved in Parkinsonian tremor 2,3 and deep brain stimulation of elements in this circuit have been shown to reduce the severity of Parkinsonian tremor4.
Prior neuroimaging studies have found have altered cerebellar activation during motor execution, and motor learning5,6, suggesting that activation in the cerebellum may reflect PD-related impairment. This cerebellar activation may be a compensatory mechanism for abnormal activity in the basal ganglia. Insight into cerebellar functional network alterations in PD may yield new targets for deep brain stimulation, and lead to improved treatment of parkinsonian symptoms. In this work, we use resting-state functional MRI (fMRI) to ascertain connectivity changes in the cerebellum from PD.
Methods
A cohort consisting of 30 subjects (15 PD and 15 age-matched control subjects) were scanned in this study. All subjects gave written, informed consent. Subject demographic data is summarized in Table 1.
Data were acquired on a 3 T MRI scanner (Prisma Fit, Siemens Medical Solutions, Malvern, PA) using a 64-channel receive only coil. MP-RAGE images (echo time (TE)/repetition time (TR)/inversion time=3.02/2600/800 ms, flip angle (FA)=8°, voxel size=0.8×0.8×0.8 mm3) were used for registration from subject space to common space.
Resting-state fMRI data were collected with a gradient echo EPI sequence using parameters matched to the HCP Lifespan project: TE/TR = 37.0/750 ms, FA=56 degrees, FOV = 216 × 188 mm2, matrix size of 108×94, slice thickness=2 mm, 64 slices, AP phase encoding, and multiband acceleration factor=8. A pair of images from a spin echo EPI sequence was acquired with imaging parameters matched to the gradient echo EPI and opposite phase encoding directions (AP and PA phase encodes) was collected to derive field maps for correction of susceptibility-related distortions in the gradient echo EPI images.
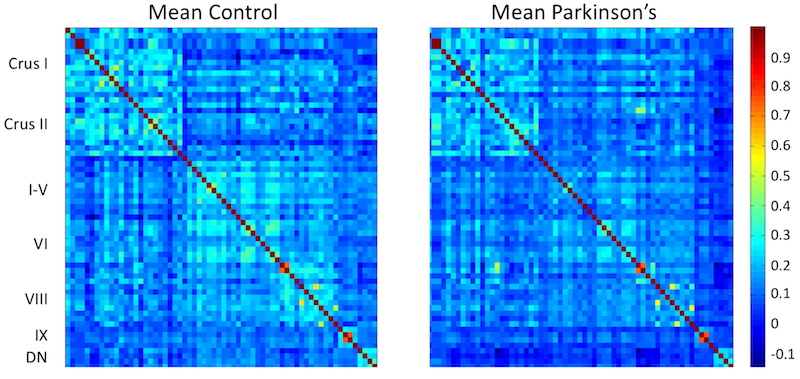
Resting-state fMRI data were preprocessed with corrections for slice timing, motion, and susceptibility, spatial smoothing using a Gaussian kernel with full width half max of 2, and CSF signal regression, and subsequently transformed to Montreal Neurological Institute (MNI) 152 space. Sixty regions of interest (ROIs) were chosen in the cerebellar lobules and 4 ROIs were chosen in the dentate nucleus. Dentate nucleus was localized using a previously published atlas7 and cerebellar lobules were localized using the cerebellar atlas in MNI152 space included in FSL. Time series within each ROI was averaged and band-pass filtered, and used to derive correlations between ROIs.
Results and Discussion
Reduced functional connectivity was observed between ROIs in lobule V and ROIs in lobules I-VI, VIII, and IX as well as with the dentate nucleus. Reduced functional connectivity was found between dentate nucleus and all ROIs in the cerebellar cortex. No difference in functional connectivity was observed within lobule VII (Crus I and Crus II). Reduced connectivity was seen within the ROIs in lobule VIII.
Prior work found motor activation from a finger tapping task in lobules V and VIII6. Reduced functional connectivity within lobules V and VIII observed in the present study may be related to Parkinsonian tremor. Further, the observation of a reduction in dentate nucleus functional connectivity with the cerebellar cortex may be in line with previously observed correlation of iron deposition in dentate nucleus with Parkinsonian tremor3. Interestingly, no differences in functional connectivity were observed within lobule VII (Crus I and Crus II), which is reported to be involved with language tasks6.
Acknowledgements
This work was supported by the Michael J. Fox Foundation (MJF 10854) and NIH-NINDS (1K23NS105944-01A1).References
[1] Wu and Hallett. The cerebellum in Parkinson’s disease. Brain 136:696–709.
[2] Guehl, et al. Tremor-related activity of neurons in the ‘motor’ thalamus: changes in firing rate and pattern in the MPTP vervet model of parkinsonism. Eur J Neurosci 17:2388-400.
[3]. He, et al. Dentate nucleus iron deposition is a potential biomarker for tremor-dominant Parkinson’s disease. NMR Biomed 30:e3554.
[4] Coenen et al. Modulation of the cerebello-thalamo-cortical network in thalamic deep brain stimulation for tremor: a diffusion tensor imaging study. Neurosurgery 75: 657–669.
[5] Wu, et al. Regional homogeneity changes in patients with Parkinson’s disease. Hum Brain Mapp 30:0:1502–10.
[6] Stoodley, Functional topography of the cerebellum for motor and cognitive tasks: An fMRI study. Neuroimage 59:1560-1570.
[7] He, et al. Improved Neuroimaging Atlas of the Dentate Nucleus. Cerebellum 6:951-956.
Figures