2556
MRI assessment of glymphatic function in the non-human primate brain1Advanced Imaging Research Center, Oregon Health & Science University, Portland, OR, United States, 2Division of Neuroscience, Oregon National Primate Research Center, Beaverton, OR, United States, 3Surgery, Oregon National Primate Research Center, Beaverton, OR, United States, 4Diagnostic Radiology, Oregon Health & Science University, Portland, OR, United States, 5Anesthesiology, Oregon Health & Science University, Portland, OR, United States
Synopsis
The astrocyte mediated exchange of cerebrospinal fluid and interstitial fluid comprise the glymphatic system, a physiology that facilitates waste removal in the brain parenchyma. Impaired solute and waste clearance may contribute to neurodegenerative conditions, and may also be associated with age. Here, we present preliminary measurements of glymphatic function in healthy adult and aged rhesus macaque brain via intrathecal injection and DCE-MRI. We demonstrate that kinetics of GBCA distribution in the CNS occur on timescales amenable to study using DCE-MRI techniques. Our preliminary results indicate that impairment in glymphatic physiology occurs with age in the rhesus macaque.
Introduction
The astrocyte mediated exchange of cerebrospinal fluid (CSF) and interstitial fluid (ISF) comprise the glymphatic system, a physiology that facilitates waste removal in the brain parenchyma.1 Impaired solute and waste clearance may contribute to neurodegenerative conditions, and may also be associated with age. Recent work has explored glymphatic function in rat brain via serial magnetic resonance imaging before and after intrathecal injection of gadolinium based contrast agent (GBCA).2 Here, we present preliminary measurements of glymphatic function in healthy adult and aged rhesus macaque brain via intrathecal injection and DCE-MRI.Methods
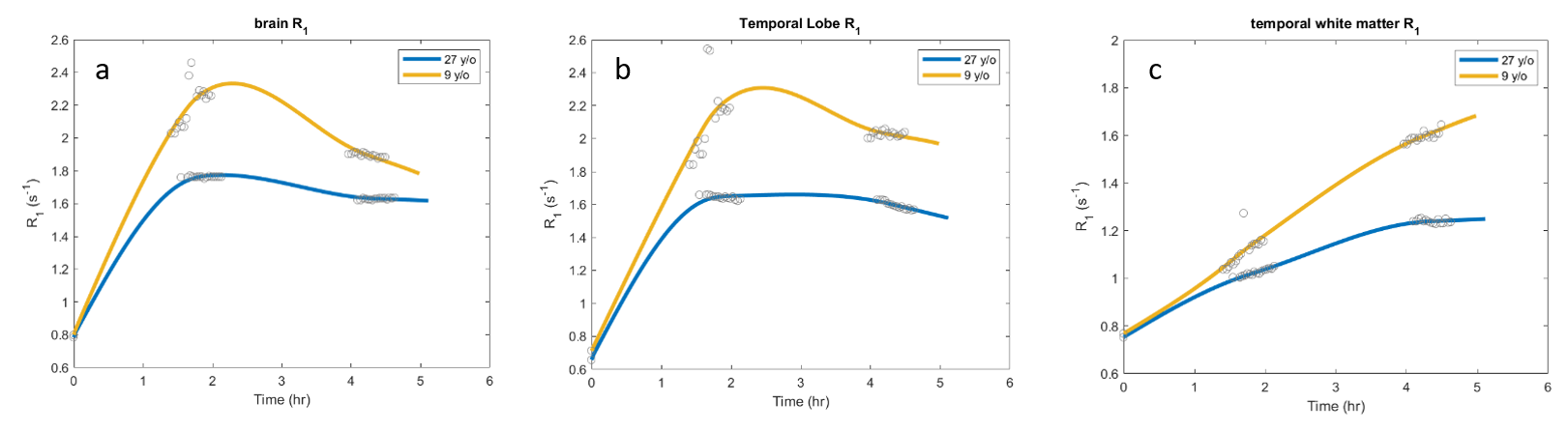
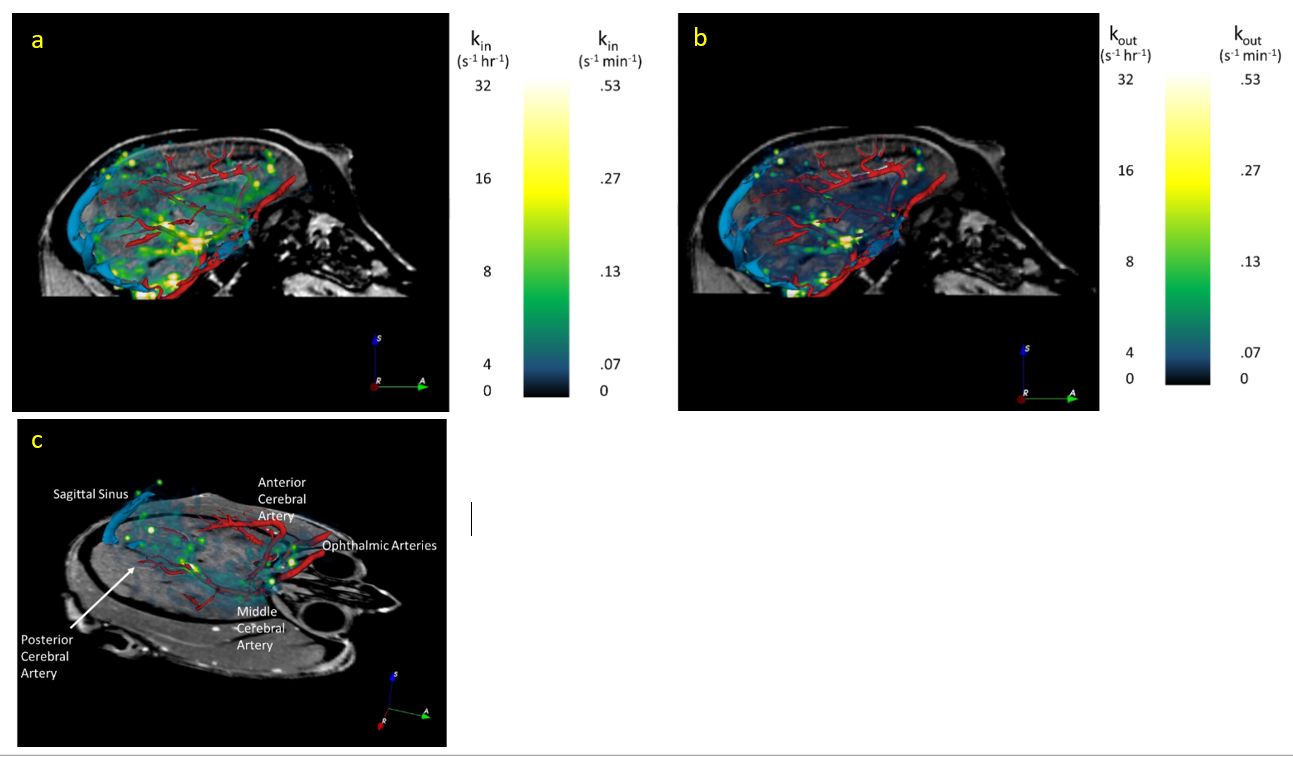
Three male rhesus macaques (age 6, 9, and 27 y/o) were included in the study. All MRI data were acquired on a whole-body Siemens Prisma 3 T MRI instrument (Erlangen, Germany) using a quadrature radiofrequency (RF) pediatric head/neck receive coil. Animals were initially sedated with Telazol, intubated and maintained on 1% isoflurane in 100% O2 and were continuously monitored by pulse oximetry, respiration, and end tidal CO2 levels during the study. Quantitative R1 (≡1/T1) mapping was performed via variable flip angle (VFA) 3D gradient recalled echo (GRE) with four flip angles (FA; 5°, 10°, 20°, 30°; TR 9ms/TE 1.7ms; 1mm isotropic resolution; (192mm)2 x 128mm field of view (FOV); 2 min 5 sec per FA). Baseline anatomical and R1 images were acquired, followed by intrathecal injection of 100µL GBCA (Gadoteridol; 2x diluted in saline) outside of the MRI. One animal (6 y/o) was returned to the MRI immediately after intrathecal injection to observe kinetics during the first 3 hrs after GBCA administration. The other two animals were imaged in 1 hr intervals for up to 6.5 hrs post-intrathecal injection; anesthesia was maintained with isoflurane outside of the MRI suite between imaging sessions. DCE data were acquired using an identical GRE acquisition with FA 20°, collected in 30 minute intervals (15 volumes). VFA R1 maps were collected before each DCE acquisition for the two animals imaged at 1 hr intervals. All DCE data for each animal were coregistered to the baseline GRE (FSL FLIRT), and corrected for motion (AFNI 3dvolreg). VFA R1 maps were then used to convert coregistered, motion-corrected DCE data to R1 maps. DCE R1 data were temporally smoothed and interpolated using a voxelwise spline fit (MATLAB); kin and kout maps (time average dR1/dt during wash-in and wash-out, respectively) were calculated using the spline fit for each voxel or for anatomic regions of interest defined by a modified version of the INIA19 rhesus brain atlas (see Figures 1 and 2).3 MR angiography was separately obtained in the 6 y/o animal: time-of-flight angiograms (FA 18°, TR/TE 22/4.31 ms, 181mm x 200mm FOV, 384 x 331 matrix, 0.5mm slice thickness) were collected pre- and post-IV injection of 0.2mmol/kg GBCA. A 3D vascular model created using 3DSlicer (v4.6.2); coregistered kin and kout maps were overlaid to assess association between glymphatic function and vascular territories.Results
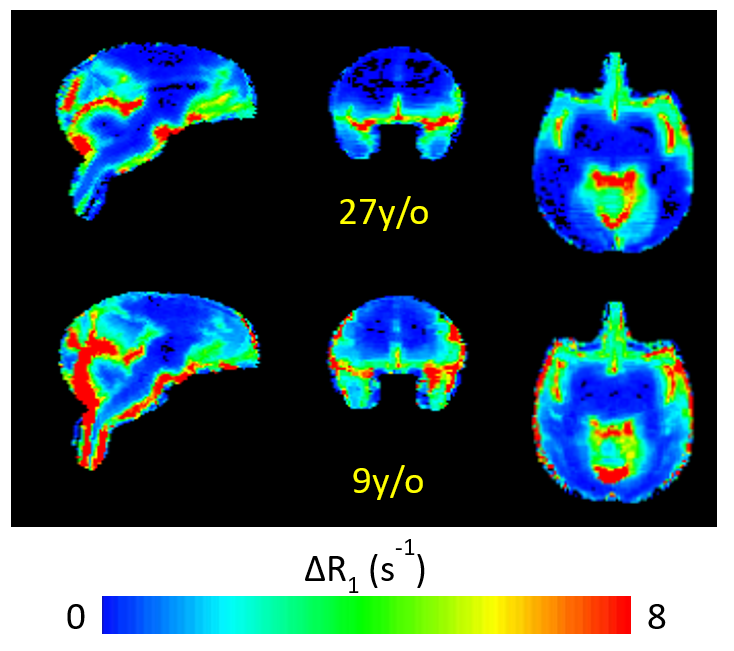
CSF-ISF interchange was reduced in the aged animal compared to the younger animals both in terms of rate of transport (kin) and maximum R1 enhancement visualized in parametric maps and by region-of-interest time course plots. Efficiency of transport, estimated by kin, appears greatest in close proximity to large vessels, particularly around the middle cerebral arteries (Figure 2). Contrast enhancement was primarily observed in the cortical gray matter, with only mild enhancement in subcortical white matter; thus, kout could not be estimated in subcortical white matter. ΔR1 maps (Figure 3) illustrate increased transport of GBCA through cortex and into cerebral white matter, indicating improved glymphatic function, in the 9 y/o animal compared to the 27 y/o.Discussion
While the rodent brain has been valuable to appreciate fundamental aspects of CSF-ISF interchange, the overall size and morphological differences (gyri and sulci, gray-matter white matter spatial distributions) between rodent and human brain are substantial and limit the generalization of findings to human brain. The non-human primate brain is closer in size and morphology to human brain, and represents a more relevant system to investigate complex fluid dynamics characteristic of the glymphatic physiology. Here, we demonstrate that kinetics of GBCA distribution in the CNS occur on timescales amenable to study using DCE-MRI techniques. We investigated the GBCA dynamics in young and old adult rhesus macaque brain. Our preliminary results indicate that impairment in glymphatic physiology occurs with age in the rhesus macaque.Acknowledgements
ONPRC P51 OD011092, Paul G Allen Frontiers Group, NIH (S10 RR027694), W. M. Keck Foundation, and Oregon Opportunity Fund.References
1. Iliff, J. J. et al. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci. Transl. Med. 4, 147ra111 (2012).
2. Benvensite, H. et al. Anesthesia with Dexmedetomidine and Low-dose Isoflurane Increases Solute Transport. Anesthesiology 127, 976–988 (2017).
3. Rohlfing, T. et al. The INIA19 template and NeuroMaps atlas for primate brain image parcellation and spatial normalization. Front. Neuroinform. 6, 718–724 (2012).
Figures