2548
Effects of Transcranial Direct Current Stimulation on Metabolite Concentrations in Children1Department of Radiology, University of Calgary, Calgary, AB, Canada, 2Alberta Children's Hospital Research Institute (ACHRI), Calgary, AB, Canada, 3Hotchkiss Brain Institute, Calgary, AB, Canada, 4Child and Adolescent Imaging Research (CAIR) Program, Calgary, AB, Canada, 5Department of Neuroscience, University of Calgary, Calgary, AB, Canada, 6Calgary Pediatric Stroke Program, Alberta Children's Hospital, Calgary, AB, Canada, 7Department of Pediatrics, University of Calgary, Calgary, AB, Canada, 8Department of Psychiatry, University of Calgary, Calgary, AB, Canada, 9The Mathison Centre for Mental Health Research and Education, University of Calgary, Calgary, AB, Canada, 10Strategic Clinical Network for Addictions and Mental Health, Calgary, AB, Canada
Synopsis
Transcranial direct current stimulation (tDCS) is a form of non-invasive brain stimulation that safely modulates brain activity. Several studies have shown that tDCS of the motor cortex facilitates motor learning and plasticity but there is little information on the underlying mechanisms. This analysis of metabolite changes in response to 1mA tDCS using typical PRESS and MEGA-PRESS is important in developing a complete understanding of the effects of stimulation. In this pediatric study, we did not detect the same GABA and glutamate changes in response to tDCS that have been seen in the adult literature.
Introduction
Transcranial direct current stimulation (tDCS) is a form of non-invasive brain stimulation in which a weak electrical current is passed between two electrodes placed on the scalp. This causes a shift in cortical excitability, for example when the primary motor cortex (M1) is targeted with the anode, cortical excitability increases1. Previous research has shown that tDCS can improve symptoms in several neurological disorder including migraine, depression, stroke, Parkinson’s as well as pain disorders with minimal side effects2. TDCS appears to enhance motor learning in pediatric populations and increases in motor skills were retained at 6 weeks after the tDCS intervention3,4. Such studies support the use of tDCS as a therapeutic tool in children but the biological mechanisms remain unknown. Adult studies show motor learning to be associated with a decrease in GABA and an increase in glutamate in M15. Pharmacological studies suggest that cortical changes induced by tDCS are associated with long-term potentiation that is mediated by glutamatergic and GABAergic neurons6. With anodal tDCS, the tissue targeted by stimulation shows GABA decreases and glutamate increases 7-10. These metabolite changes have been associated with changes in task performance7. It is unknown if these findings translate to a pediatric population and how long changes in metabolites persist.
Methods
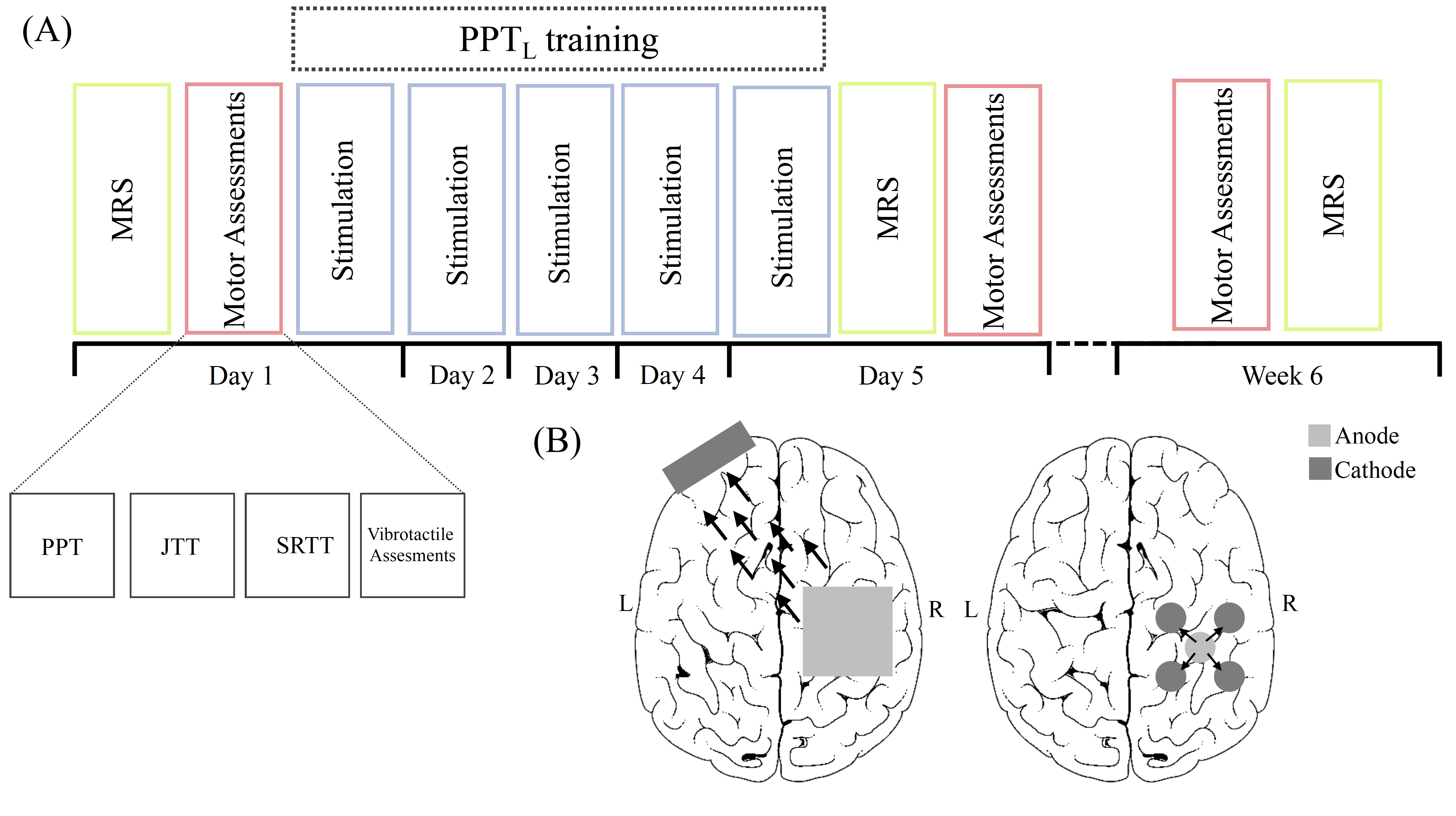
Twenty-four typically developing, right-handed children (age 12 – 18 years) participated in five consecutive days of tDCS intervention targeting the right M1 while training in a motor task (Purdue Pegboard Task) with their left hand (Fig 1A). Participants were randomly assigned to one of three experimental groups (Fig 1B):
- Anodal tDCS – the conventional application of tDCS
- High Definition tDCS – a recent innovated with improved localization of stimulation
- Sham tDCS – a negative control condition
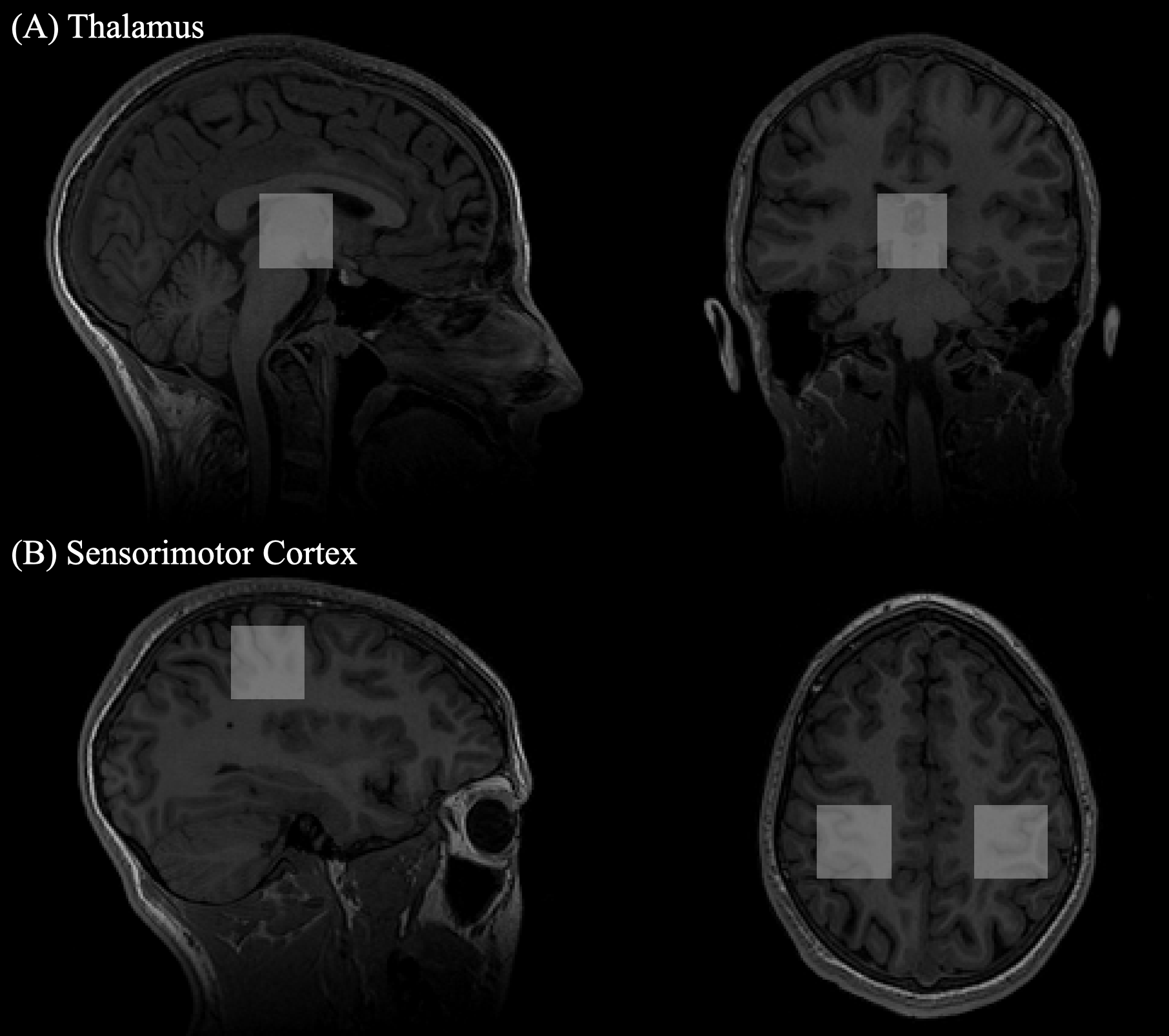
Spectroscopy data was collected on a 3T GE MRI scanner at baseline, after the 5 days of tDCS and 6-weeks later. A 32-channel head coil was used. Metabolite levels were measured in 30×30×30 mm3 voxels in both sensorimotor cortices and the thalamus, placed anatomically (Fig 2). The sensorimotor cortices are involved in acquisition of motor skill while the thalamus has been identified as a relay center for motor function. Glutamate was quantified using a conventional PRESS sequence (TR/TE = 1800/35 ms, 64 averages with 8 unsuppressed water scans) and quantified using LC Model11 followed by CSF-tissue correction. GABA was measured using a MEGA-PRESS sequence12 (TR/TE = 1800/68 ms, 256 averages with 14 ms editing pulses applied at 1.9 ppm and 7.46 ppm alternating every two averages and 16 unsuppressed water scans) and quantified using GANNET13, including tissue correction to account for increased concentration of GABA in grey matter14. Changes in GABA and glutamate between tDCS conditions and over time were assessed using a repeated measures ANOVA analysis with age and sex covariates for each voxel and a Tukey Test was used to correct for multiple comparisons in follow-up paired analyses.
Results
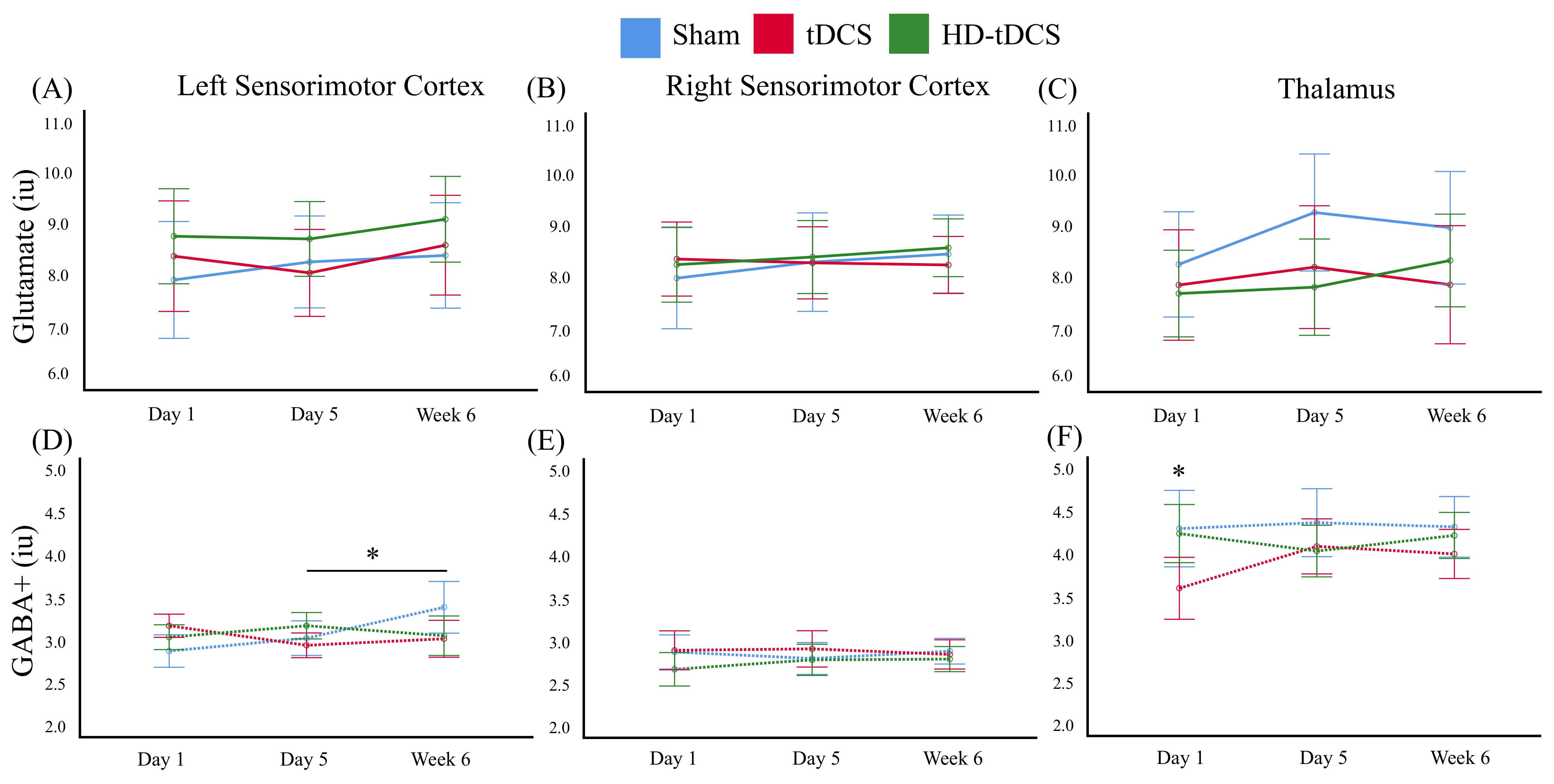
A trend toward increased glutamate was observed in the thalamus after the 5 days of intervention (p = 0.054) in the sham condition (Figure 3C). No other glutamate changes were seen.
In the left M1, increased GABA was seen between day 5 of intervention and 6 weeks follow up (p = 0.015) in the sham condition (Figure 3D). No other significant changes were detected between time points.
Discussion
This is the first study that we are aware of that explores tDCS induced metabolite changes in a pediatric population. While adult studies have shown anodal tDCS increases M1 glutamate and decrease GABA7 , this study suggests the developing brain responds to tDCS differently. The absence of glutamate changes in both the right and left M1 over time differs from the current adult literature that suggests glutamate increases following tDCS targetting M1. This could potentially be attributed to the fact that the developing brain is in a continual state of learning and therefore GABA and glutamate are already at a respectively decreased and increased state to support this dynamic brain state. The increase in GABA between day 5 and 6 week follow up in the left M1 was unexpected and may reflect GABA equilibrating upon consolidation of a skill that is enhanced by transcallosal inhibition. Due to our small sample size, further investigation is required before conclusive interpretations can be made.Conclusion
The current study showed no alteration in metabolite concentrations following 20 minutes of various montages of tDCS. These results suggest pediatric populations show different metabolite responses to tDCS compared to adult literature necessitating additional studies.Acknowledgements
Study participant recruited from the University of Calgary Healthy Infants and Children's Clinical Research Program (HICCUP) and research was performed in collaboration with the Non-Invasive Neurostimulation Network (N3) of the University of Calgary.
References
- Reis J, Fritsch B. Modulation of motor performance and motor learning by transcranial direct current stimulation. Curr Opin Neurol. 2011;24(6):590–6.
- Nitsche MA, Cohen LG, Wassermann EM, Priori A, Lang N, Antal A, et al. Transcranial Direct Current Stimulation: State of the art 2008. Brain Stimulation. 2018; 1(3); 206-23.
- Ciechanski, P., Kirton, A. Transcranial Direct-Current Stimulation Can Enhance Motor Learning in Children. Cerebral Cortex. 2017;27(5), 2758-2767.
- Cole L, Giuffre A, Ciechanski P, Carlson HL, Zewdie E, Kuo H, Kirton, A. Effects of High-Definition and Conventional Transcranial Direct-Current Stimulation on Motor Learning in Children. 2018;12(October):1–12.
- Floyer-Lea A. Rapid Modulation of GABA Concentration in Human Sensorimotor Cortex During Motor Learning. J Neurophysiology. 2006; 95(3): 1639-44.
- Froc DJ, Chapman A, Trepel C, Racine RJ. Long-Term Depression and Depotentiation in the Sensorimotor Cortex of the Freely Moving Rat. J Neuroscience, 2000; 20(1): 438-45.
- Stagg CJ, Best JG, Stephenson MC, O’Shea J, Wylezinska M, Kincses ZT, et al. Polarity-Sensitive Modulation of Cortical Neurotransmitters by Transcranial Stimulation. J Neurosci. 2009;29(16):5202–6.
- Stagg CJ, Bachtiar V, Johansen-Berg H. The role of GABA in human motor learning. Curr Biol. 2011;21(6):480–4.
- Stagg CJ, Jayaram G, Pastor D, Kincses ZT, Matthews PM, Johansen-Berg H. Polarity and timing-dependent effects of transcranial direct current stimulation in explicit motor learning. Neuropsychologia. 2011;49(5):800–4.
- Clark VP, Coffman BA, Trumbo MC, Gasparovic C. Transcranial direct current stimulation (tDCS) produces localized and specific alterations in neurochemistry: A 1H magnetic resonance spectroscopy study. Neurosci Lett. 2011;500(1):67–71.
- Provencher SW. Estimation of metabolite concentrations from localized in vivo proton NMR spectra. Magn Reson Med. 1993;30(6):672–9.
- Mescher M, Merkle H, Kirsch J, Garwood M, Gruetter R. Simultaneous in vivo spectral editing and water suppression. NMR in Biomedicine. 198; 11(6):266-72.
- Edden RAE, Puts NAJ, Harris AD, Barker PB, Evans CJ. Gannet: A batch-processing tool for the quantitative analysis of gamma-aminobutyric acid-edited MR spectroscopy spectra. J Magn Reson Imaging. 2014;40(6):1445–52.
- Harris AD, Puts NAJ, Edden RAE. Tissue correction for GABA-edited MRS: Considerations of voxel composition, tissue segmentation, and tissue relaxations. J Magn Reson Imaging. 2015;42(5):1431–40.
Figures