2507
Metabolic fate of glucose in atypical meningioma studied by 13C NMR isotopomer analysis1Neurosurgery, Houston Methodist Hospital and Research Institute, Houston, TX, United States
Synopsis
Meningiomas are tumors arising from meninges, the membranes surrounding the brain and the spinal cord. Majority of meningiomas (grade-1) are benign and grow slowly. However, atypical meningiomas (grade-2) exhibit increased cellular abnormalities, and grow at a faster rate than benign meningiomas. Moreover, atypical meningiomas prone to recurrence and show resistance to radiotherapy. Atypical meningiomas show higher 18F-FDG uptake in PET scans. No prior reports are available on investigating metabolic fate of glucose in atypical meningiomas. The goal of this study is to probe the metabolic fate of glucose using NMR based [U-13C]glucose isotopic tracing methods in patient-derived atypical meningioma cells.
Introduction
Meningiomas are the most frequently diagnosed CNS tumors that arise from meningeal membranes surrounding the brain and spinal cord. They represent about 30% of all CNS tumors. World Health Organization (WHO) classifies meningiomas into three types based on histology [1,2]. Grade I meningiomas are benign growths and are slow growing. Grade-II, known as “atypical” meningiomas represent 10-15% and show high index of recurrence. Grade III meningiomas are anaplastic malignancies and represent about 1-3% of meningiomas. Atypical menigiomas exhibit high uptake of 18F-FDG by PET imaging. Lactate and alanine levels are higher in atypical meningiomas indicating altered glucose metabolism similar to malignant brain tumors [3,4]. However, metabolic fate of glucose in human atypical meningiomas is largely unknown. In this study, we investigate [U-13C]glucose metabolism in a patient-derived atypical meningioma cell line using NMR spectroscopy and isotopomer analysis.Methods
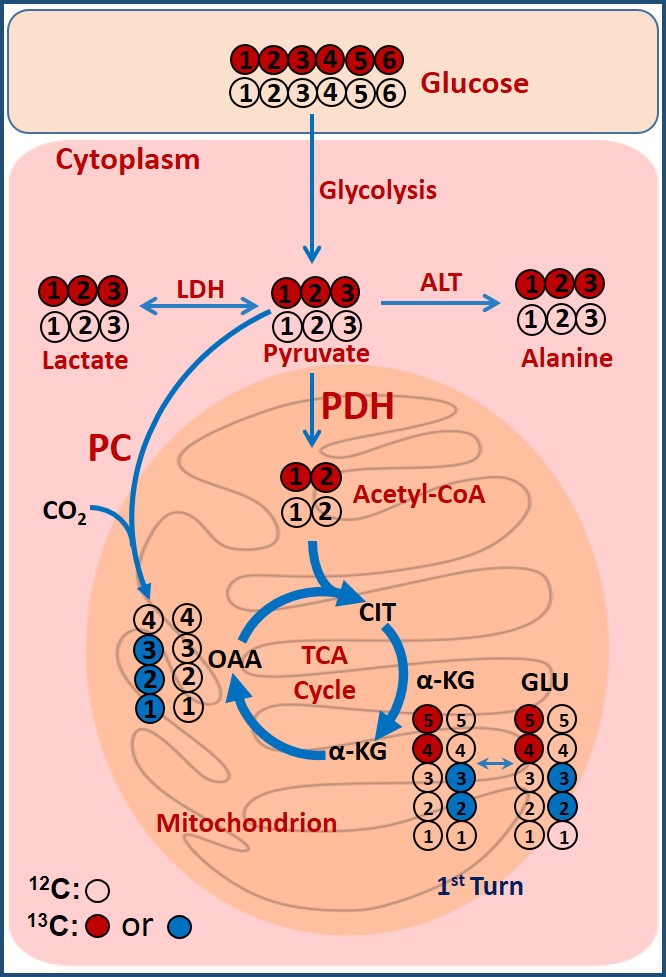
Tumor tissues were collected from atypical meningioma patients undergoing craniotomy at the Houston Methodist Hospital following an approved IRB protocol. Cells were isolated from the tumor tissue and were initially grown in ABMTM Astrocyte Basal Medium, incubated at 37°C under humidified air with 5% CO2. During the final 6 hours, 15.0 mM [U-13C]glucose was introduced to the glucose-free Dulbecco’s modified Eagle’s medium (DMEM). After 6 hours, the medium was removed, cells were washed with PBS buffer and the cell pellets were snap-frozen in liquid nitrogen. Prior to NMR data collection, the cell pellets were extracted in 5% perchloric acid, extracts were lyophilized and were reconstituted in 180 µL D2O containing 1.0 mM DSS-d6 (pH = 7.4). The sample-solutions were transferred to a 3-mm NMR tubes, and 1D 1H and 1H-decoupled 13C NMR spectra were collected on a Bruker 600 MHz spectrometer equipped with a 5-mm cryo-probe optimized for direct 13C detection.Discussion
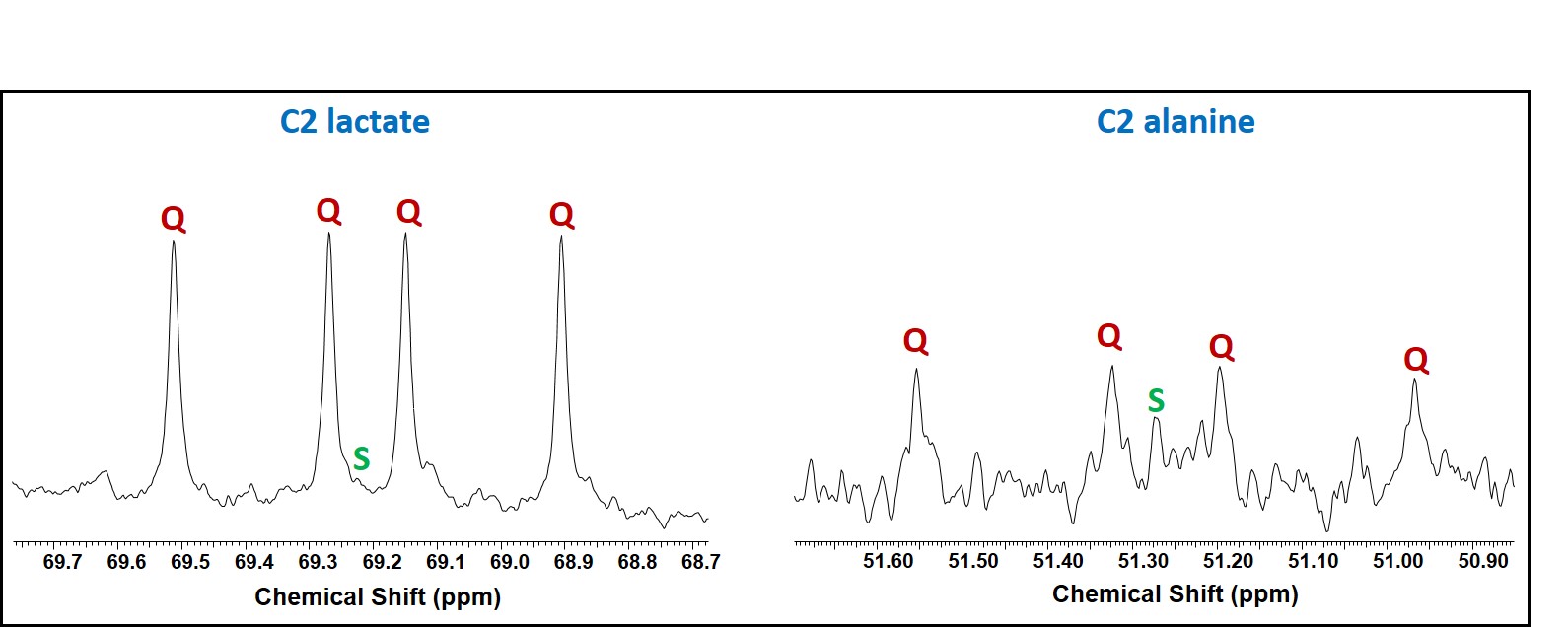
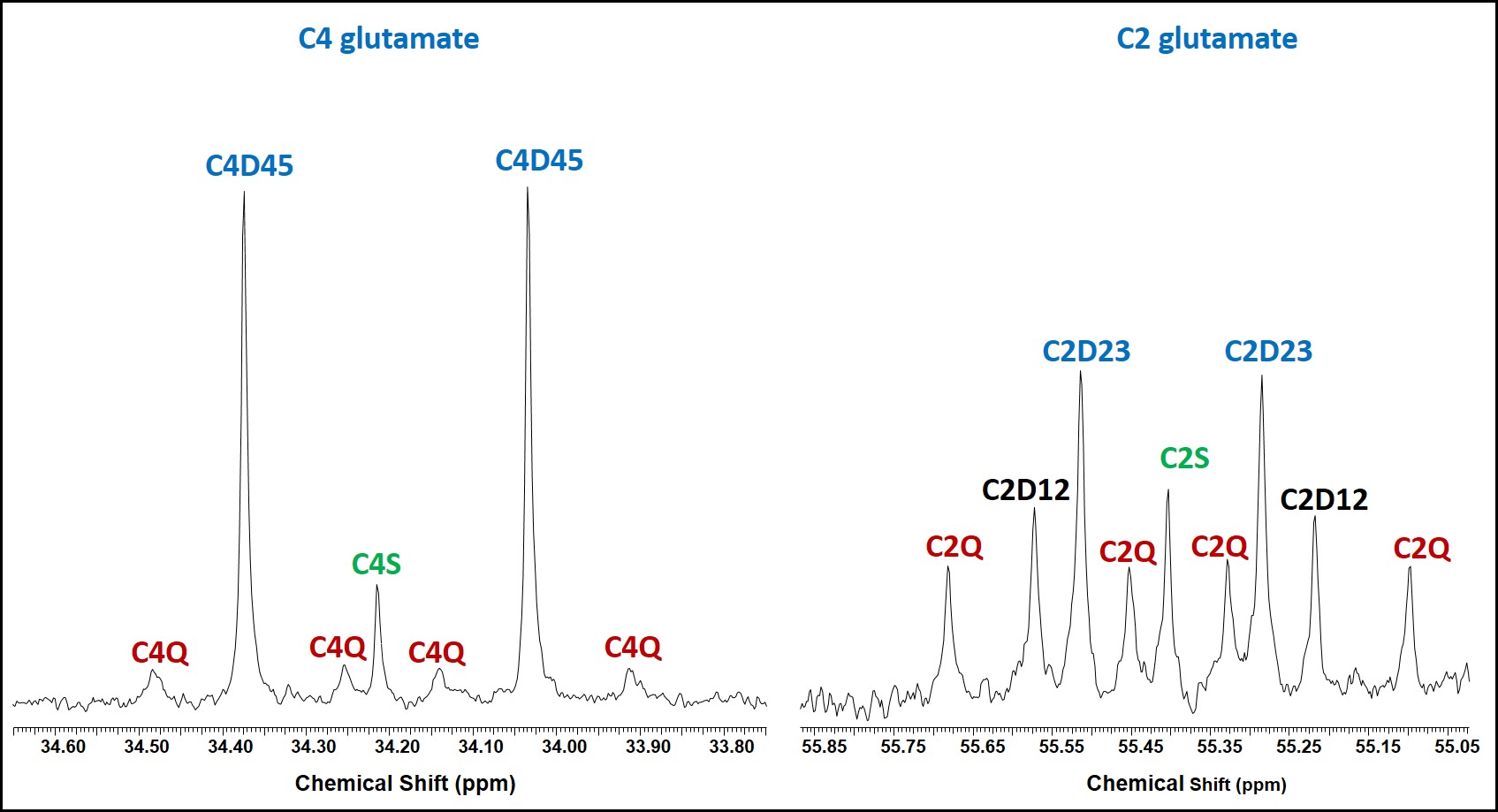
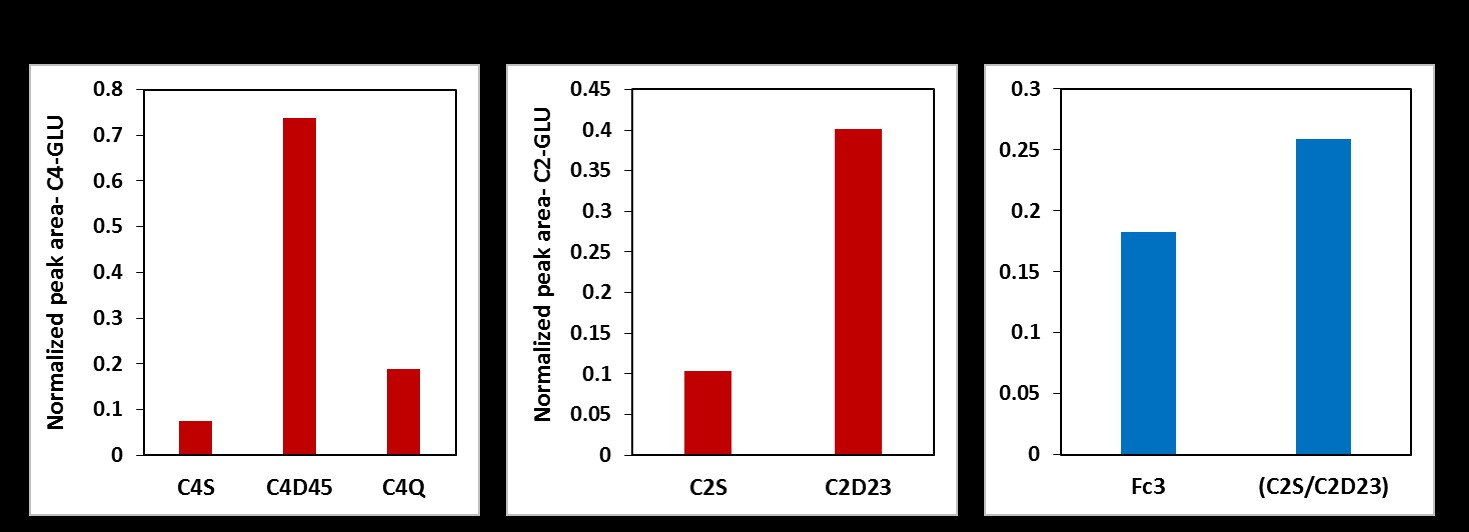
Two major findings in this study are: First, in addition to producing lactate, atypical meningioma cells are capable of oxidizing glucose in the TCA cycle to generate energy via acetyl-CoA production through PDH activity. Second, these cells show active anaplerosis through the presence of PC activity. Proliferating cells need high energy to produce carbon precursors/intermediates to synthesize macromolecules that are generated within the TCA cycle in the mitochondria. The flux through PC in these cells replenishes OAA in order to maintain continuous flow of TCA cycle intermediates for anabolic processes [5].Acknowledgements
We thank Peak Center for financial support. We thank Blessy John and Sophie Lopez for technical assistance.
References
Monleon D, Morales JM, Gonzales-Darder J et al., Benign and atypical meningioma metabolic signatures by HR-MAS molecular profiling. J Proteome Res. 7 (2008) 2882-2888.
Monleón D, Morales JM, Gonzalez-Segura A, Gonzalez-Darder JM, Gil-Benso R, Cerdá-Nicolás M, López-Ginés C. Metabolic aggressiveness in benign meningiomas with chromosomal instabilities. Cancer Res. 70 (2010) 8426-8434.
E.A. Maher, I. Marin-Valencia, R.M. Bachoo, T. Mashimo, J. Raisanen, K.J. Hatanpaa, A. Jindal, F.M. Jeffrey, C. Choi, C. Madden, D. Mathews, J.M. Pascual, B.E. Mickey, C.R. Malloy, R.J. DeBerardinis, Metabolism of [U-13 C]glucose in human brain tumors in vivo, NMR. Biomed. 25 (2012) 1234-1244.
K. Pichumani, T. Mashimo, H-M. Baek, J. Ratnakar, B. Mickey, R.J. DeBerardinis, E.A. Maher, R.M. Bachoo, C.R. Malloy, Z. Kovacs, Conditions for 13C NMR detection of 2-Hydroxyglutarate in tissue extracts from isocitrate dehydrogenase-mutated gliomas, Anal. Biochem. 481 (2015) 4-6.
Cheng T, Sudderth J, Yang C, Mullen AR, Jin ES, Matés JM, DeBerardinis RJ. Pyruvate carboxylase is required for glutamine-independent growth of tumor cells. Proc Natl Acad Sci U S A. 108 (2011) 8674-8679.
Figures