2279
Brain regional susceptibility to Oxidative Stress in a rat model of Chronic Hepatic Encephalopathy: in-vivo 1H MRS, ex-vivo ESR spectroscopy and histology findings1Center for Biomedical Imaging, EPFL, Lausanne, Switzerland, 2Service of Biomedicine, CHUV, Lausanne, Switzerland, 3Swiss Center for Liver Disease in Children, Department of Pediatrics, HUG, Lausanne, Switzerland, 4Laboratory of Physics of Complex Matter, EPFL, Lausanne, Switzerland, 5Swiss Center for Liver Disease in Children, Department of Pediatrics, HUG, Geneva, Switzerland
Synopsis
Oxidative stress (OS) is thought to be an important factor in chronic hepatic encephalopathy (CHE) pathogenesis. Combining in-vivo 1H-MRS, ex-vivo ESR and histology of CNS allowed us to investigate the course and brain regional difference of cerebral OS in the rat model of CHE. Early changes in brain antioxidants (Asc and GSH, post-BDL) were validated by ESR and histology. Our results showed a stronger vulnerability of cerebellum and confirmed that the antioxidant system impairment and central OS is an early event in CHE.
PURPOSE
Oxidative stress (OS) is believed to be an important feature in the pathogenesis of chronic hepatic encephalopathy (CHE)1 and is thought to contribute to inflammation. In CHE impaired ammonium clearance by the diseased liver will lead to brain glutamine accumulation. In vitro, impaired ammonium detoxification together with glutamine increase induces reactive oxygen species (ROS) generation associated with astrocyte impairment2,3. ROS play an important role in cellular signalling, synaptic plasticity, learning and memory. When in excess or when antioxidant defense systems are weakened, they cause cellular damage4,5. We have previously shown the indirect presence of OS as a decrease of brain Asc in the hippocampus of a rat model of CHE. We have further validated these findings using for the first time ESR spectroscopy6. How different brain regions will react to CHE and will OS show a brain regional vulnerability are still unknown. Therefore, in the present study, we focused on the evolution of two brain antioxidants, Asc and GSH in cerebellum using in-vivo longitudinal 1H-MRS followed by ex-vivo ESR detection of ROS and histological measures to assess the OS levels and potential brain regional differences. Cerebellum data were compared to previously acquired hippocampus data6.METHODS
In-vivo 1H-MRS: 9.4T-MR-system(Varian/Magnex-Scientific),a home-built 14mm-diameter quadrature 1H-surface coil as a transceiver and the SPECIAL sequence7 were used (TE=2.8ms,TR=4sec;160averages). Cerebellum (2.5×2.5×2.5mm3) of Wistar male adult rats was scanned before bile-duct-ligation (BDL-animal model of CHE) and after every 2weeks up to week8 (n=6). Metabolites concentrations were calculated by LCModel using water as reference.
Ex vivo ESR: ESP300E(Bruker BioSpin,TE102 cavity) spectrometer was used for intracellular O2.-quantification. Cerebellum was extracted at 6weeks post-BDL/sham-surgery from additional rats (n=6), weighed, sliced and transferred into RPMI1640 medium with 10mM CMH cell-permeable non-toxic spin-trap (Noxygen GmbH) and incubated at 37oC. After each incubation time, tissue suspension was transferred into quartz capillary and measured.
Staining: Hippocampus/cerebellum were incubated in the dark (37oC, 60min) with Nitro Blue Tetrazolium (1.6mg/mL) for histochemical detection of ROS. Immunohistochemical staining using GPX1(anti-oxidative enzyme) and GFAP(astrocyte intermediate filament) on 10mm OCT-embedded brain sections was performed.
RESULTS AND DISCUSSION
Increase in plasma NH4+ and bilirubin confirmed the presence of chronic liver disease.
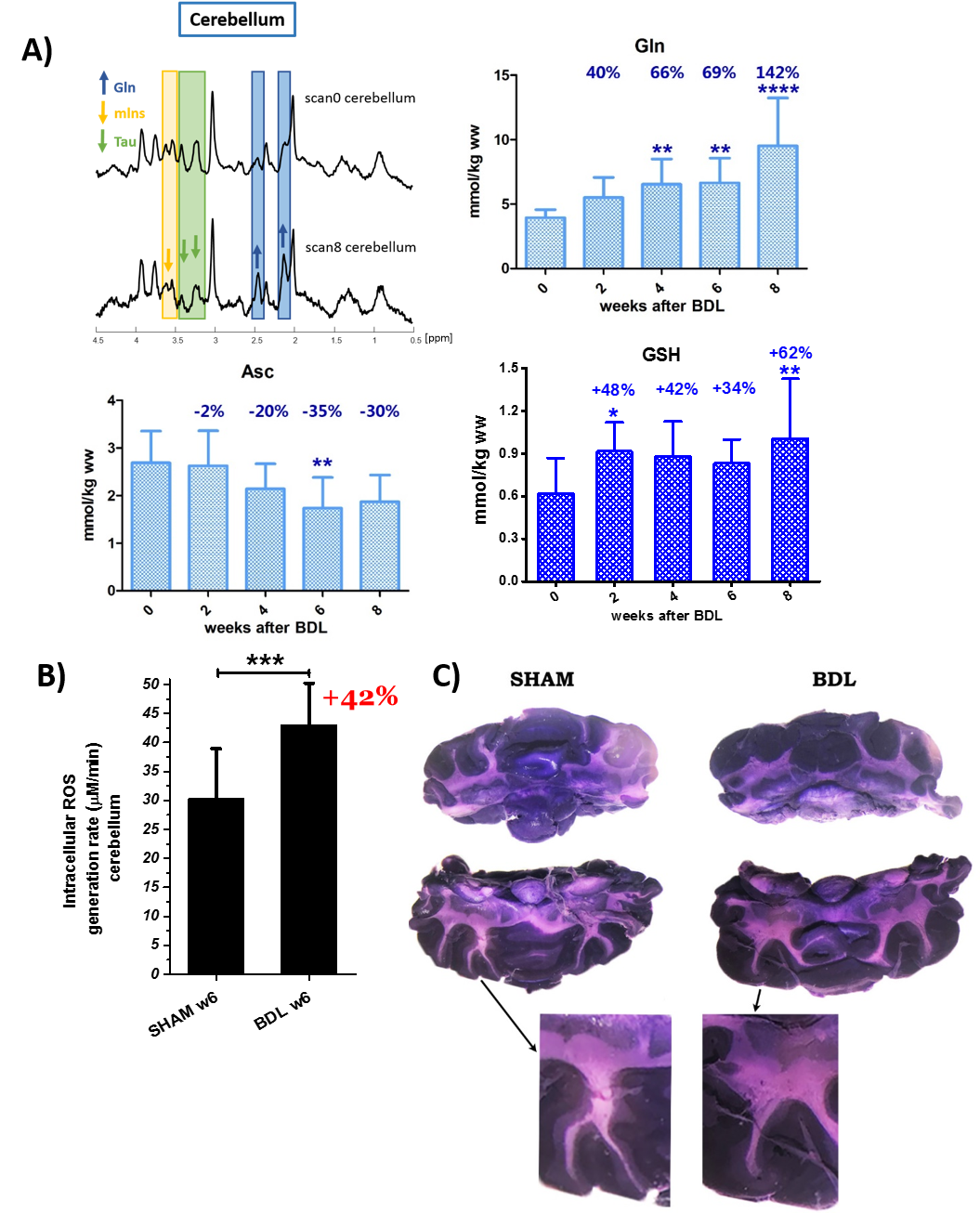
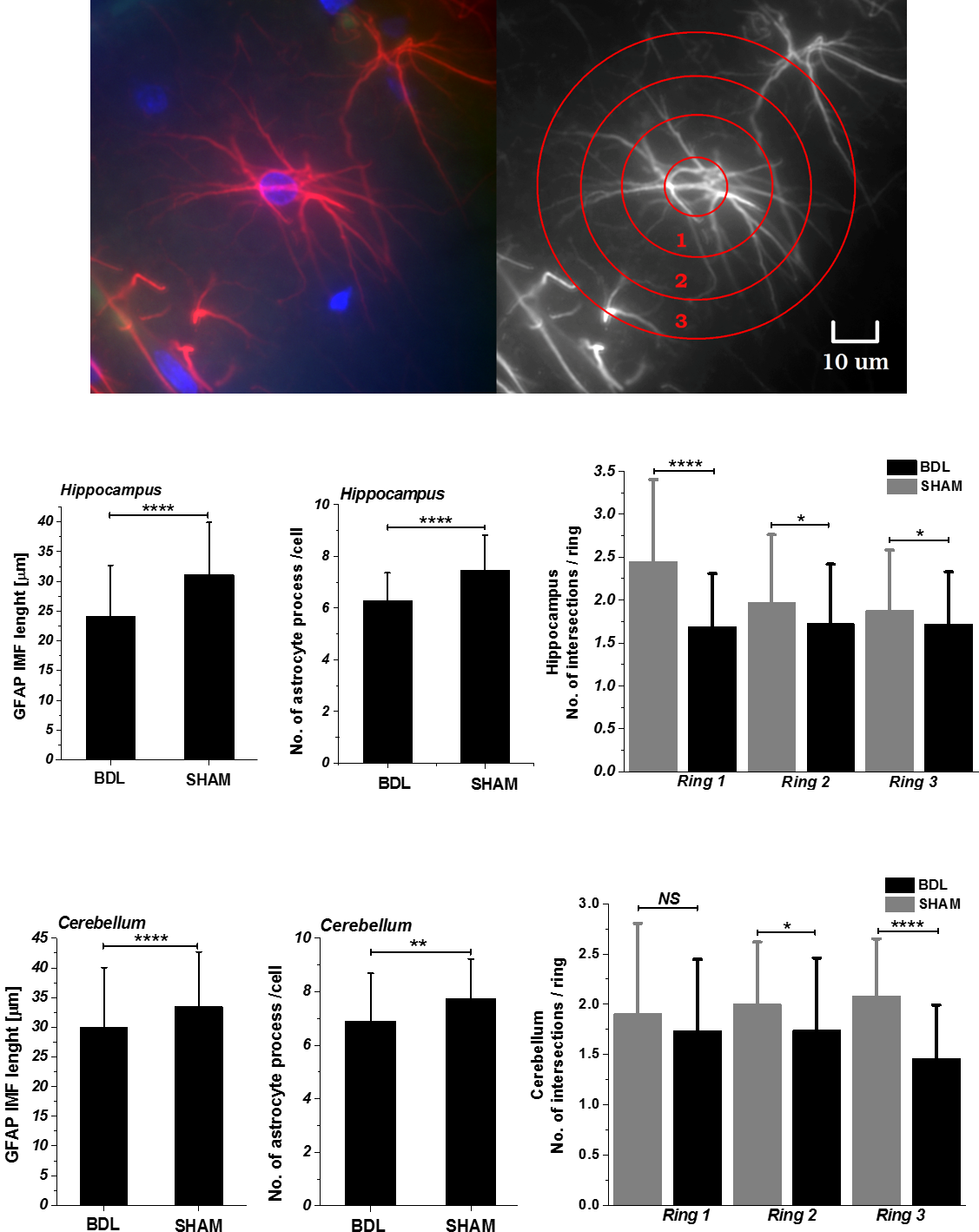
1H-MRS showed a differential brain regional vulnerability to CHE. The cerebellum showed the stronger increase in glutamine (+142% vs +107% hippocampus6) and the corresponding osmotic answer (stronger decrease in main osmolytes: Ins,Tau). Glutamine, a byproduct of ammonia metabolism has a toxic effect by mimicking ammonia and producing the OS response in cells8. Moreover, astrocytes are essential to detoxify ammonium9. Cytoskeleton plays a key role in OS responses10, therefore accumulation of intracellular ROS can lead to astrocytes intermediate filaments degeneration. An overall atrophy of astrocyte processes length and intersections number in BDL’s rats was observed (Fig.1) suggesting an astrocytic activation and synaptic depression induction11.
The two important brain antioxidants (Asc,GSH) also showed a differential regional response. Asc showed a stronger decrease in cerebellum reaching the maximal decrease at week8 (hippocampus -15%,p<0.0166 vs cerebellum -35%,p<0.01)(Fig.2A). GSH had a different behavior: an increase in cerebellum (+62%-8weeks post-BDL,p<0.01) vs a trend of decrease in hippocampus6.
Immunohistochemical staining with GPX1-enzyme protecting from oxidative damage12,13, revealed an increase activity in Purkinje and granular cell layer of cerebellum in BDL rats (Fig.3) but not in hippocampus. Purkinje cells of BDL rats also showed shrinking soma with mean size 8.04±1.8mm2 vs 12.5±1.2mm2 in shams (p<0.00001). Increased GPX1 in cerebellum suggests that GSH synthesis may increase in response to OS related injury.
Ex-vivo ESR results revealed a significant difference in redox state between the two brain regions (~31%,p<0.004) in sham rats (23.15±5.14mM/min - hippocampus vs 30.31±8.62 mM/min - cerebellum). However, the relative change between BDL's and shams was similar ~42%. Significant increase of hippocampal and cerebellar OS in BDL (p<0.01,p<0.001) supported the 1H-MRS findings of decreased Asc concentrations (Fig.2AB).
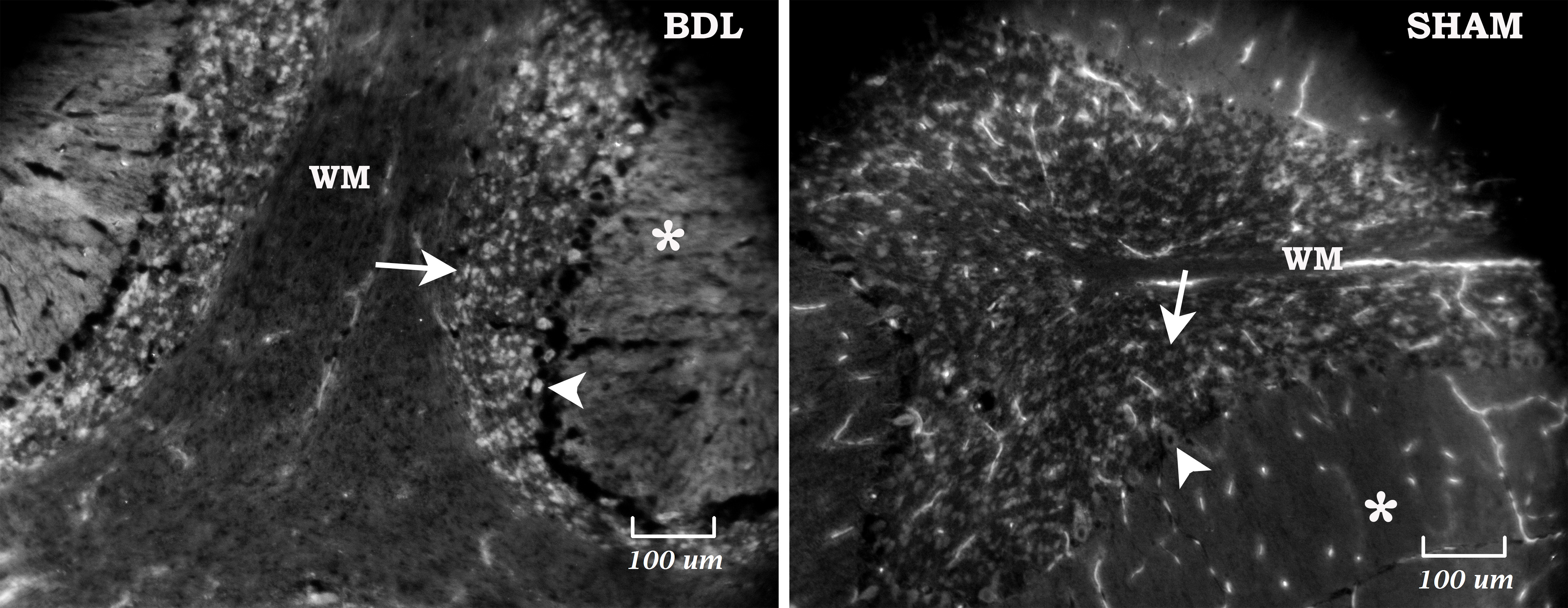
Increased ROS in white matter (WM) of hippocampus-fimbria hippocampi6 and cerebellum in BDL rats was detected by NBT staining (Fig.2C). Elevated OS in WM tracts might suggest that ROS plays an important role in brain functions disruption14-16.
CONCLUSIONS
Our results indicate a different susceptibility of cerebellum and hippocampus to CHE, reflected mainly in brain Gln and antioxidants/OS, and a relationship between increased OS and interrupted antioxidant defence system. In addition, it is plausible that redox dysregulation, decreased antioxidants levels might lead to reduced WM integrity. The GFAP depicted astroglial atrophy may cause reduction in network connectivity, synaptic coverage and homeostatic capabilities17. OS is involved in the propagation of cellular injury and may be an important factor in the etiology of the CHE. Therefore, implementation of antioxidants treatment could be considered as a promising therapeutic strategy.Acknowledgements
Supported by CIBM of the UNIL, UNIGE, HUG, CHUV, EPFL, the Leenaards and Jeantet Foundations and the SNSF project no 310030_173222/1References
1Bosoi CR, Rose CF. Metab Brain Dis. 2013 Jun;28(2):175-8; 2Skowronska M. and Albrecht J., Neurochemistry International, 2012; 3Lachman V. et al., Archives of Biochemistry and Biophysics, 2013; 4Lauren T. Knapp and Eric Klann, Journal of Neuroscience Research, 2002; 5Cynthia A.et al., PNAS 2009 August, 106 (32) 13576-13581; 6K. Pierzchala et al., ISMRM2018 # 448; 7Mlynárik et al, Magn Reson Med 2006; 8Jan Albrecht and Michael D. Norenberg, HEPATOLOGY 2006,44:788-794 ; 9Gavin A. K. et al., Liver Int. 2014: 34: 1184–1191;10John Gardiner, Robyn Overall and Jan Marc, Diseases 2013, 1, 36-50;11Rogier Min & Thomas Nevian, 2012 nature NEUROSCIENENCE; 12Zhang Y. et al., Mol Vis. 2011:2325–2332. PMID: 21921984; 13Ruzhi Deng et al.,PLOS ONE | DOI:10.1371/journal.pone.0126561 May 29, 2015; 14The Hippocampus Book, Oxford University Press, 2007; 15I. O. Imosemi, Int. J. Morphol.,31(1):203-210, 2013;16 T.H. Tranah, A. Paolino, Debbie L. Shawcross, Clinical Liver Disease, Vol 5, No 3, March 2015; 17Alexei Verkhratsky1,2, José J. Rodríguez, and Luca Steardo, The Neuroscientist 2014; 18Baehner RL, Boxer LA, Blood, 1976; 19Armstrong JS et al., Int J Androl. 2002.Figures
Cerebellum: A) Longitudinal evolution of Gln, Asc and GSH, from week0 (before BDL) to 8weeks after BDL. B) ROS generation rate calculated from ESR kinetic plots (EPR signal levels versus the elapsed time). C) Cerebellum stained with NBT at week 8 after BDL: detection by histochemical staining18,19. NBT accumulation show increased ROS level in WM structure and in granular cell layer. NBT reacts specifically with O2.- forming purple formazan precipitate and can help to indirectly determine the redox status.
Data are presented as mean ± SD and statistical significance (One way Anova) is always presented compared to week 0: *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001.