2278
Evaluation of Bovine Brain Extract Phantom Structure through Polarization Transfer Solid-State NMR1Department of Biomedical Engineering, Vanderbilt University, Nashville, TN, United States, 2Vanderbilt University Institute of Imaging Science, Nashville, TN, United States, 3Physical Chemistry, Lund University, Lund, Sweden
Synopsis
Myelin MRI phantoms used to characterize magnetization dynamics require comparable microstructure and composition to physiological myelin for experiments to be translatable. Folch Fraction I bovine brain extract was evaluated for its phase behavior, composition, and structure through Polarization Transfer solid-state NMR (PT ssNMR), 31P static NMR, and small-angle X-ray scattering. Results indicate the absence of cholesterol and the coexistence of solid and liquid isotropic phases in phantoms across various solvent contents at body temperature, as opposed to physiological myelin's multi-lamellar liquid crystalline structure. Experiments aimed at characterizing myelin’s microstructure should look to other preparation methods.
Introduction
Myelin, a lamellar membrane sheath that wraps around nerve axons, significantly contributes to MRI contrast in brain tissue. Water diffusion, dipole-dipole interactions, and chemical coupling with protons on myelin lipid hydrocarbon chains all contribute to magnetization exchange and impact MRI contrast as a result, but the extent of their individual roles is not well-understood. This set of processes is difficult to parse through imaging in vivo, where physical parameters are uncontrollable and inseparable. Development of a myelin-mimetic MRI phantom will enable the characterization of magnetization exchange within myelin and the roles of specific macromolecules on acquired signal through experiments that perturb myelin’s biophysical and chemical environment. Understanding these mechanisms in greater detail will enable more specific evaluation of MRI contrast with respect to the structure and composition of brain tissue.
Recent studies on ultrashort transverse relaxation in myelin have implemented phantoms made from Folch Fraction I, a biological extract of myelin lipids found in bovine brain1,2. Extending the range of translatable experiments on this phantom to longitudinal relaxation and diffusion requires thorough knowledge of the phantom’s microstructure with respect to that of physiological myelin. Furthermore, experiments which perturb the phantom’s chemical environment necessitate exact knowledge of its composition. This experiment aims to evaluate the structure, phase behavior, and composition of phantoms made from Folch Fraction I through a series of solid-state NMR studies.
Methods
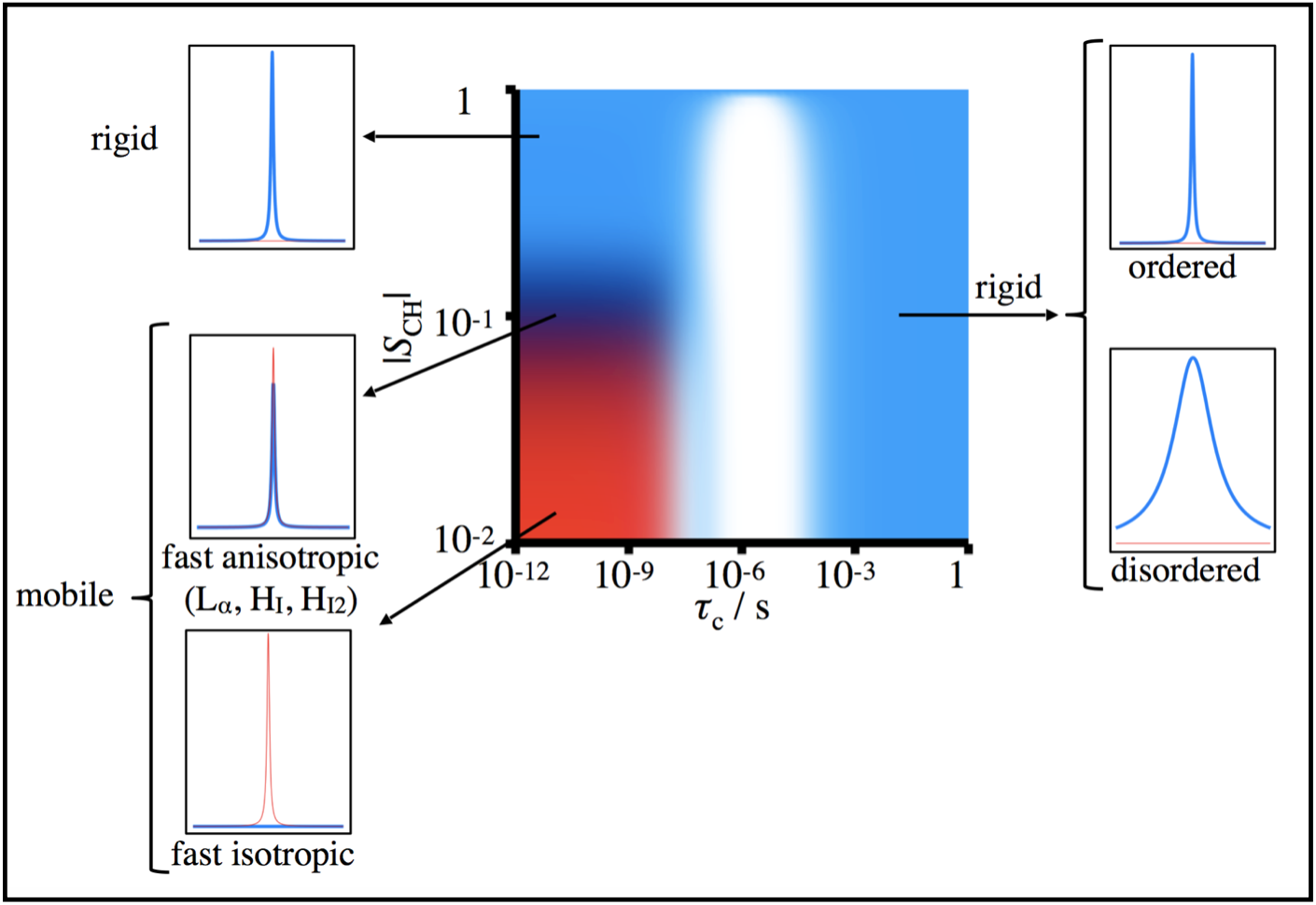
1H-13C polarization transfer solid-state NMR (PT ssNMR) compares signal amplitudes of 13C atoms obtained through cross-polarization (CP), direct polarization (DP), and insensitive nuclei enhanced by polarization transfer (INEPT) experiments to characterize systems near the solid-liquid phase border3. The selectivity of CP experiments to carbon in rigid chemical environments and of INEPT experiments to mobile carbon atoms enables simultaneous detection of solid, liquid-crystalline, and liquid isotropic components present in a given sample (see Figure 1). Once this is known, X-ray scattering can further classify the phase. For example, if PT ssNMR reveals that a myelin sample exists in the liquid-crystalline phase, X-ray scattering can validate whether that phase is lamellar or hexagonal.
A series of myelin phantoms was prepared from Folch Fraction I extract at varying solvent concentrations (deionized water to follow previous literature protocol1,2 or PBS to match physiological pH). A second batch of Folch Fraction I was dissolved with 30 wt% cholesterol in a 50:50 volumetric solution of chloroform and methanol prior to phantom preparation. CP, DP, and INEPT experiments were performed in sequence on a Bruker Avance II 500 MHz spectrometer (Lund University, Sweden). To further characterize each phantom, 1H NMR, 31P magic angle spinning (MAS) NMR, and 31P static NMR experiments were also performed on each phantom. All NMR studies were conducted at body temperature (37℃) and 5 kHz MAS unless otherwise specified. Small- and wide-angle X-ray scattering experiments were performed using a Ganesha SAXS system.
Results
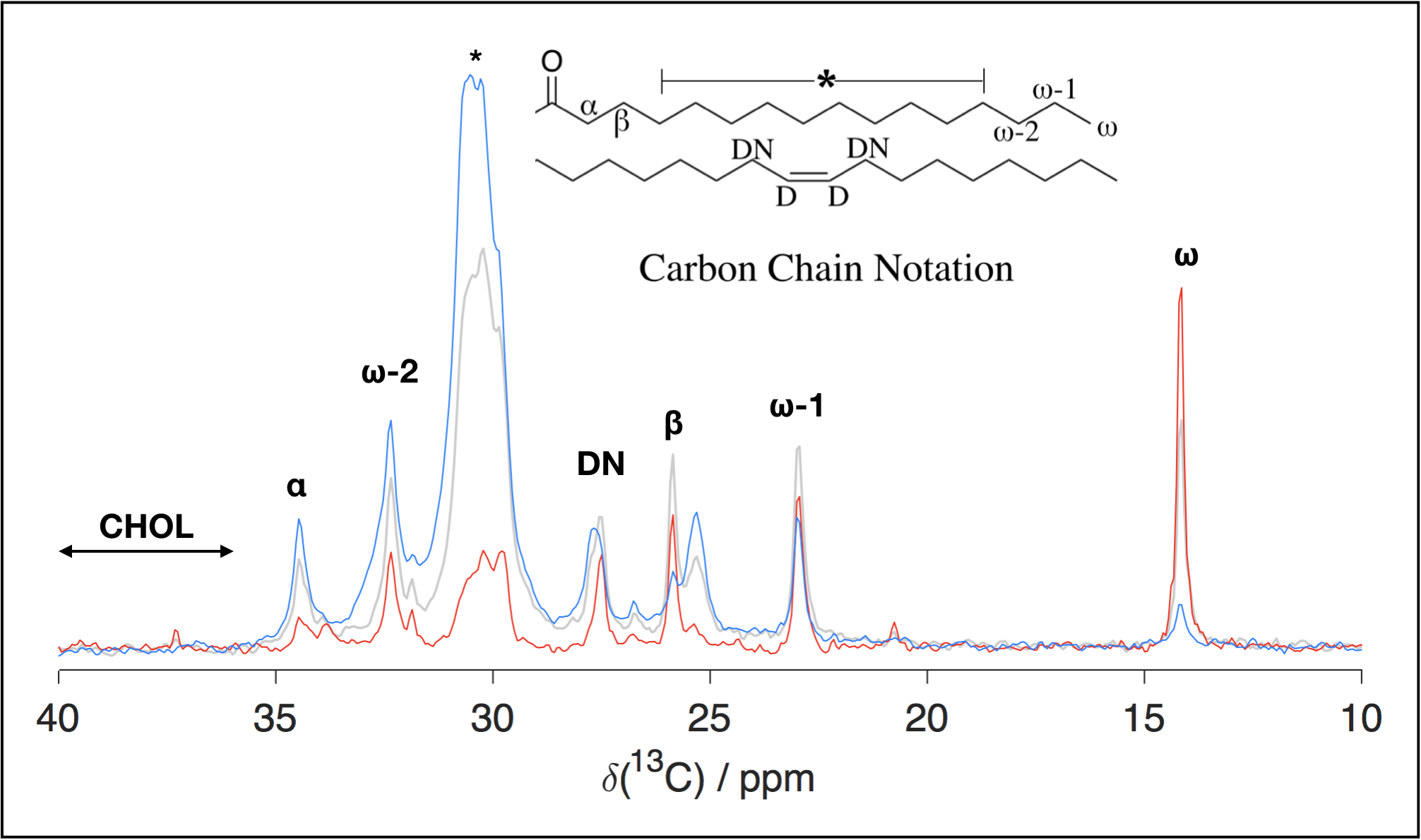
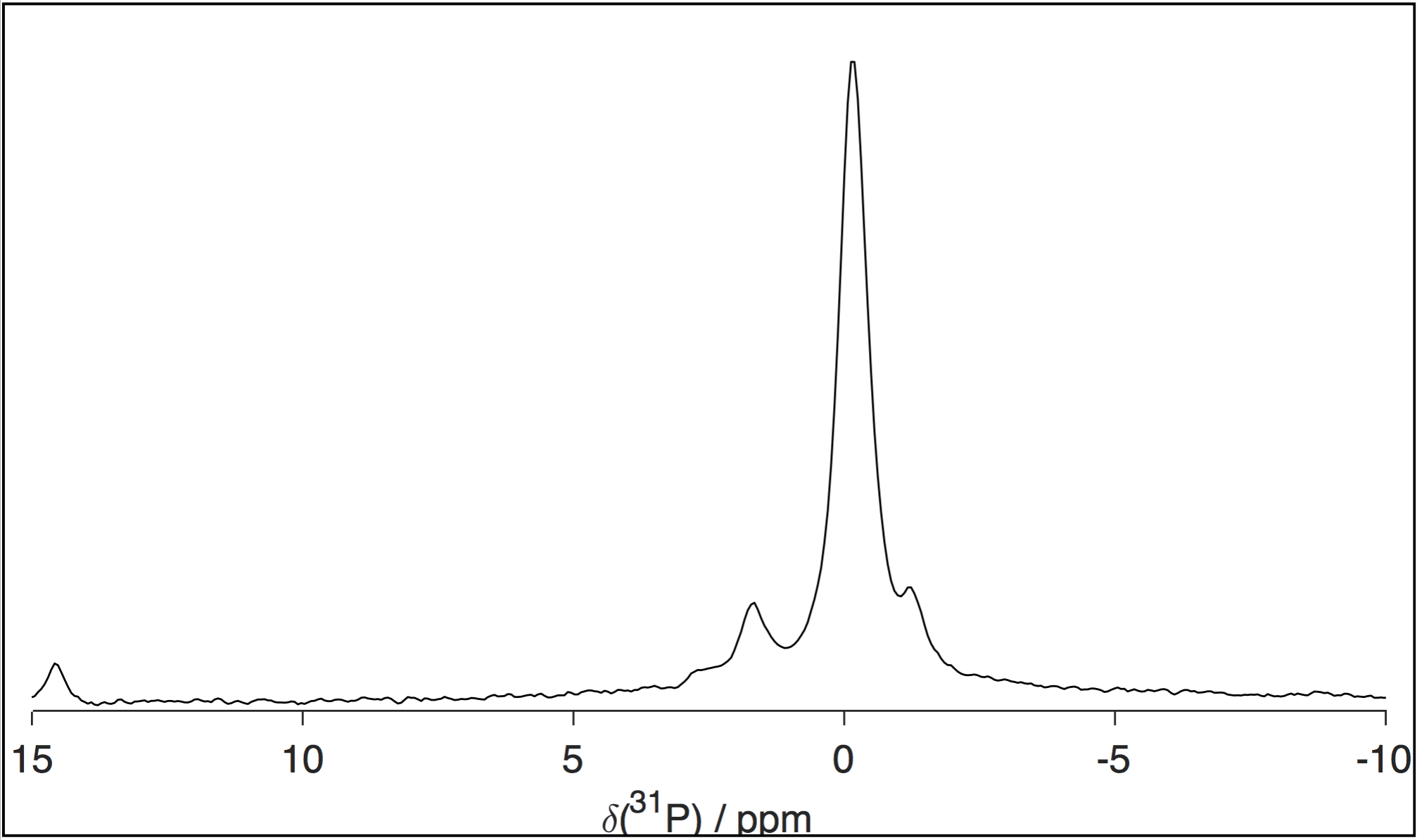
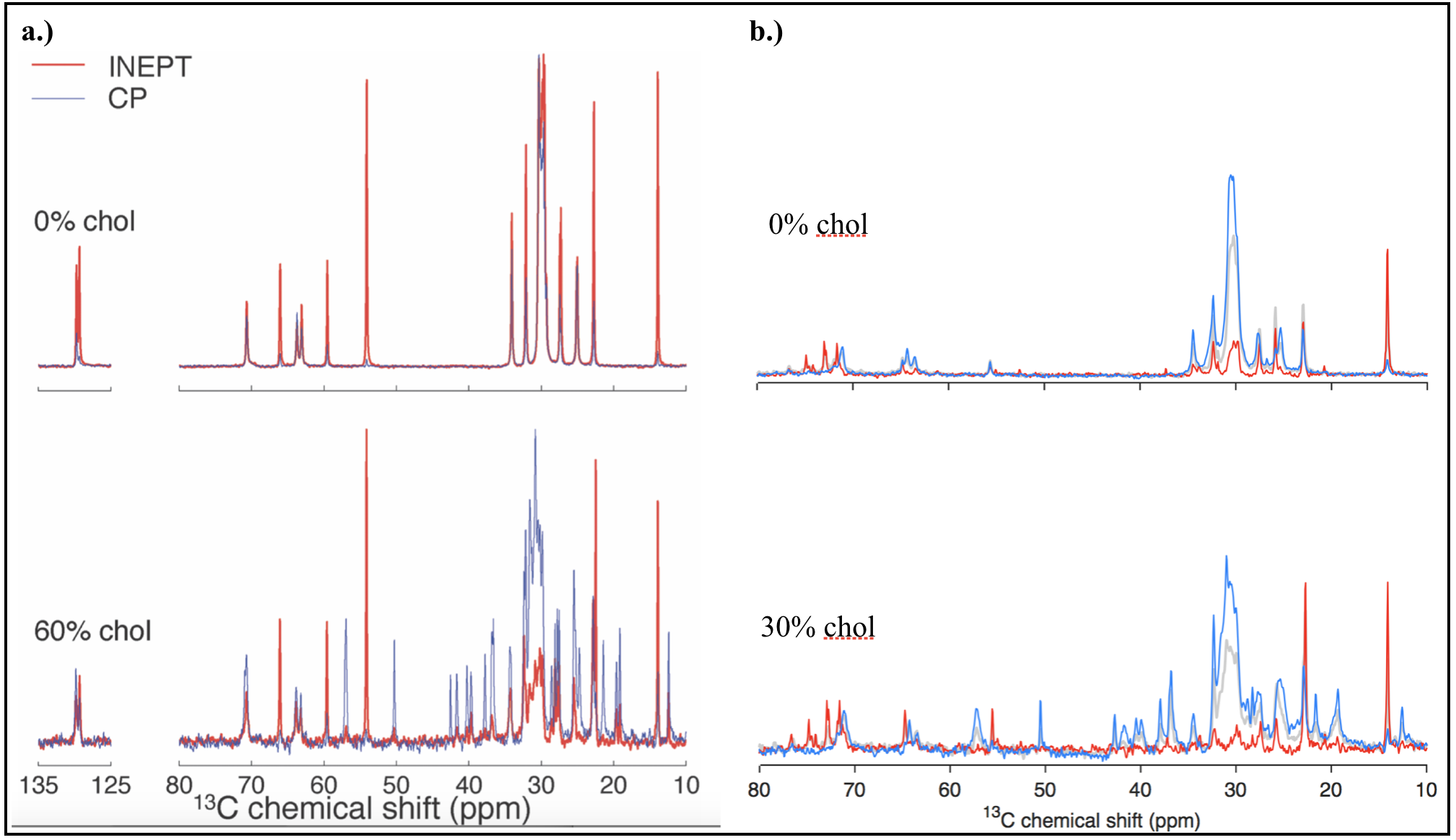
Figures 2 and 3 show, respectively, representative spectra for PT ssNMR and 31P static NMR of a phantom matching physiological myelin’s solid:liquid mass ratio. Figure 4 compares PT ssNMR spectra of the same phantom to literature spectra of liquid-crystalline phosphatidylcholine, a major constituent of myelin4. SAXS data were used to fit the phase configuration and membrane spacing of each phantom. Phantoms were modeled better by a lamellar rather than hexagonal structure; added solvent content generally decreased fitted membrane separation.Discussion
PT ssNMR experiments indicate a complete absence of cholesterol from Folch Fraction I extract, most likely a result of the fractionation process. Furthermore, while significant CP and INEPT signal are present for most peaks in PT ssNMR spectra, the signal peak shape and chemical shift of the two sequences do not match in many cases, particularly at low water content. This phenomenon persists after adding cholesterol and changing solvent, and indicates the co-existence of a solid and liquid phase rather than one liquid-crystalline phase, as would be desired in a myelin phantom. Static 31P NMR spectra display major peaks spanning only ~5 ppm in width; they lack the wide signal shelf characteristic of lamellar or hexagonal phase spectra, suggesting that phosphorous exists exclusively in an isotropic environment in these phantoms. While Folch Fraction I’s ultrashort echo time properties correlate with those of physiological myelin1, rigorous analysis through solid-state NMR reveals that its chemical structure and phase behavior are not an adequate match. Phantom studies aimed at characterizing myelin’s microstructure should look to other phantom preparation methods, including synthesis from myelin’s constituent lipids and in-lab myelin extraction from brain tissue; recent research has shown more favorable 13C and 31P spectra by directly isolating and purifying bovine brain myelin6,7.Acknowledgements
This work is financially supported by a research grant from the National Institutes of Health (NIH EB019980). Experiments conducted at Lund University in Sweden, including all NMR and SAXS experiments performed in this study, were supported by funding from the Knut and Alice Wallenberg Foundation (KAW).References
1Horch, R. A.; Gore, J. C.; Does, M. D., Magn Res Med 2011, 66 (1), 24–31.
2Sheth, V.; Shao, H.,; Chen, J.; Vandenberg, S.; Corey-Bloom, J.; Bydder, G. M.; Du, J., NeuroImage 2016, 136, 37–44.
3Nowacka, A.; Bongartz, N. A.; Ollila, O. H.; Nylander, T.; Topgaard, D., J Magn Reson 2013, 230, 165-75.
4Ferreira, T.M.; Coreta-Gomes, F.; Ollila, O.H.; Moreno, M.J.; Vaz, W.L.C.; Topgaard, D., Phys. Chem. Chem. Phys. 2013, 15, 1976-1989.
5Wilhelm, M.J.; Ong, H.H.; Wehrli, S.L.; Li, C; Tsai, P.; Hackney, D.B; Wehrli, F.W., PNAS 2012, 109(24) 9605-9610.
6Autilio, L.A.; Norton, W.T; Terry, R.D., J Neurochem 1964, 11, 17-27.
7Pham, Q.D.; Topgaard, D.; Sparr, E., PNAS 2017, 114(2) E112-E121.
Figures