2272
Characterizing the relationships between early systemic cytokine levels and neurochemical changes in Alzheimer’s disease: A longitudinal neuroimaging study in the TgF344-AD rodent model1Integrated Program in Neuroscience, McGill University, Montreal, QC, Canada, 2Douglas Mental Health University Institute, Verdun, QC, Canada, 3Department of Psychiatry, McGill University, Montreal, QC, Canada
Synopsis
Cytokines are chemical signalling molecules released by the immune system in response to pathological insults. Cytokine upregulation is an early feature of Alzheimer’s disease (AD) and a possible contributor to downstream neuropathology and cognitive decline. Using magnetic resonance spectroscopy (MRS), cytokine-specific enzyme-linked immunosorbent assays (ELISA) and behavioural measures, we aimed to investigate the relationships between cytokine activity, early neurochemical changes and cognitive decline in an AD rodent model. Preliminary metabolic alterations suggest a paradoxical increase in synaptic activity, coinciding with cognitive deficits. This ongoing study is a step towards understanding the impact of abnormal cytokine levels on the AD brain.
Introduction
Alzheimer’s disease (AD) is a progressive neurodegenerative disorder characterized by a cascade of neuropathological features, including amyloid beta (Aβ) plaques and neurofibrillary tangles (NFTs), along with cognitive decline.1 Despite massive research efforts, the causes of AD are unknown and definitive antemortem diagnosis remains elusive.
An early feature in the AD cascade is elevated systemic levels of cytokines; chemical signalling molecules released by the immune system in response to pathological insults. Cytokines interfere with neurotransmission, potentially contributing to neuronal stress, glial dysfunction and cognitive decline. Past literature notes a paradoxical increase in neuronal activity during prodromal AD, which may be linked to proinflammatory cytokines, like tumor necrosis factor alpha (TNFα).2,3 In addition, magnetic resonance spectroscopy (MRS) studies have found alterations in brain myo-inositol (mIns) and glutamate (Glu) during AD progression, representing neuronal and glial changes.4 The relationships between neurochemistry and cytokine concentrations have yet to be explored.
In this study, we investigate the relationships between early cytokine activity, neurochemistry and cognitive decline in the TgF344-AD rat model; a transgenic (Tg) model exhibiting age-dependent accumulation of Aβ and NFTs, alongside synaptic dysfunction.5 MRS is used to characterize neurochemistry, while cytokine levels are assessed using enzyme-linked immunosorbent assays (ELISA), and spatial memory and learning are addressed using the Barnes maze.5,6 MRS and ELISA measures are acquired longitudinally at Day 120, 165, 210, 255, and 300. Behavioural data is acquired at Day 120, 210 and 300.
Methods
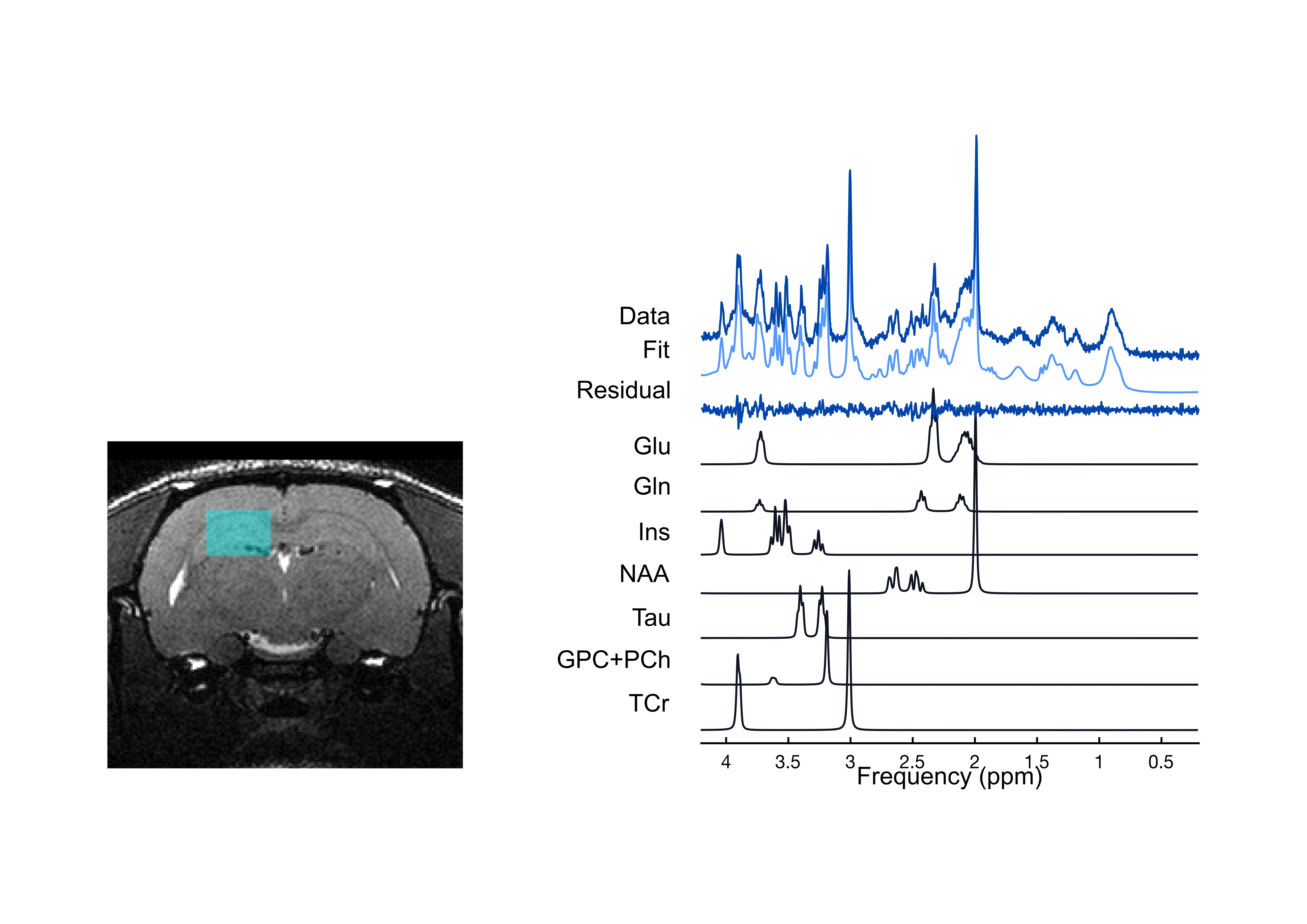
A 7 Tesla Bruker Biospec 70/30 Scanner was used to acquire localized MRS data from a 31 μL voxel (2.5 x 3.5 x 3.5) in the dorsal hippocampus (Figure 1). Shimming was performed with FASTMAP, resulting in water linewidths of 7.8 to 9.8 Hz. Localized MR spectra were acquired using the PRESS sequence (TR/TE = 3000/11). FID-A was used for pre-processing, and processed spectra were analyzed in LCModel using a basis set simulated in house using FID-A.7,8 All metabolites were referenced to total creatine.
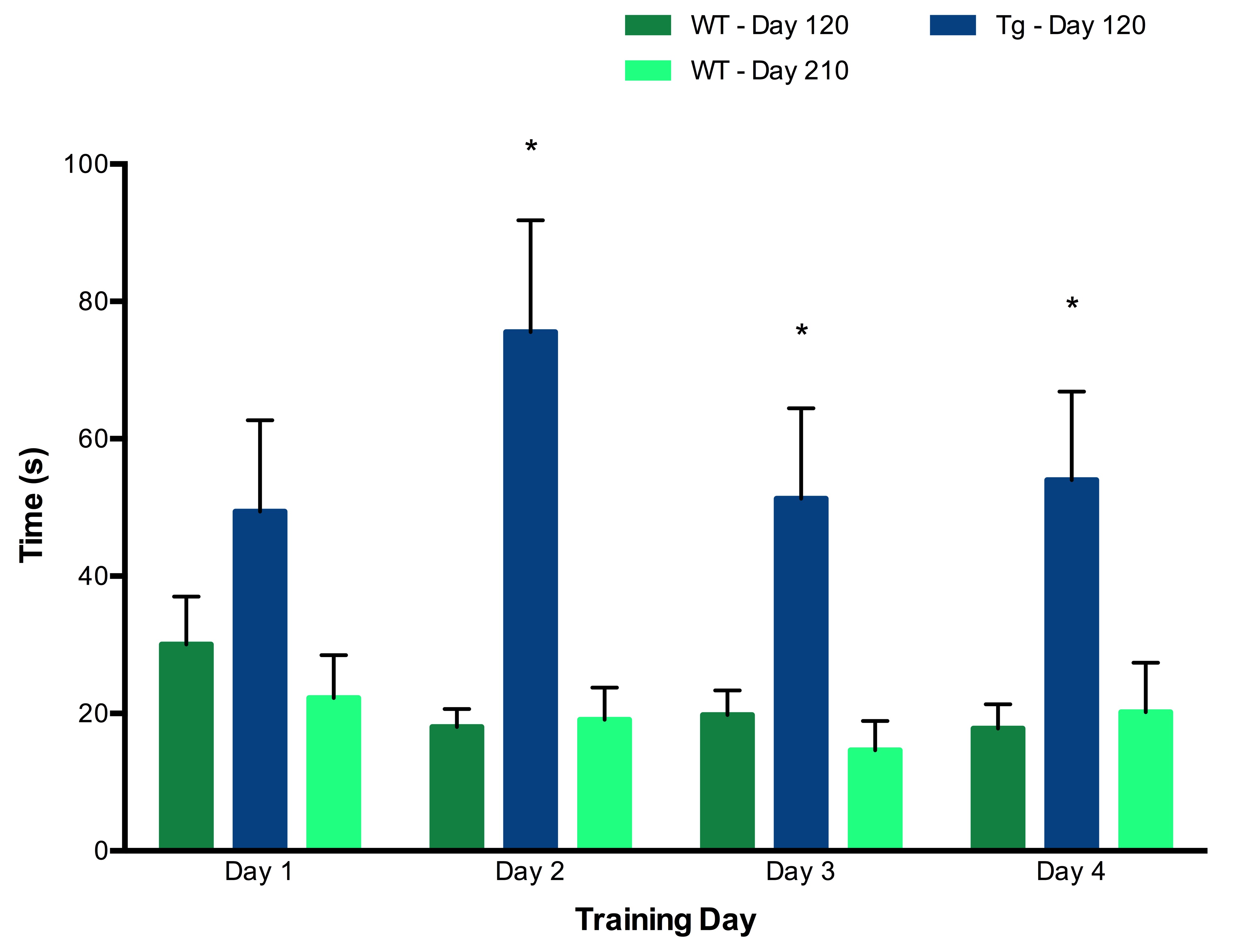
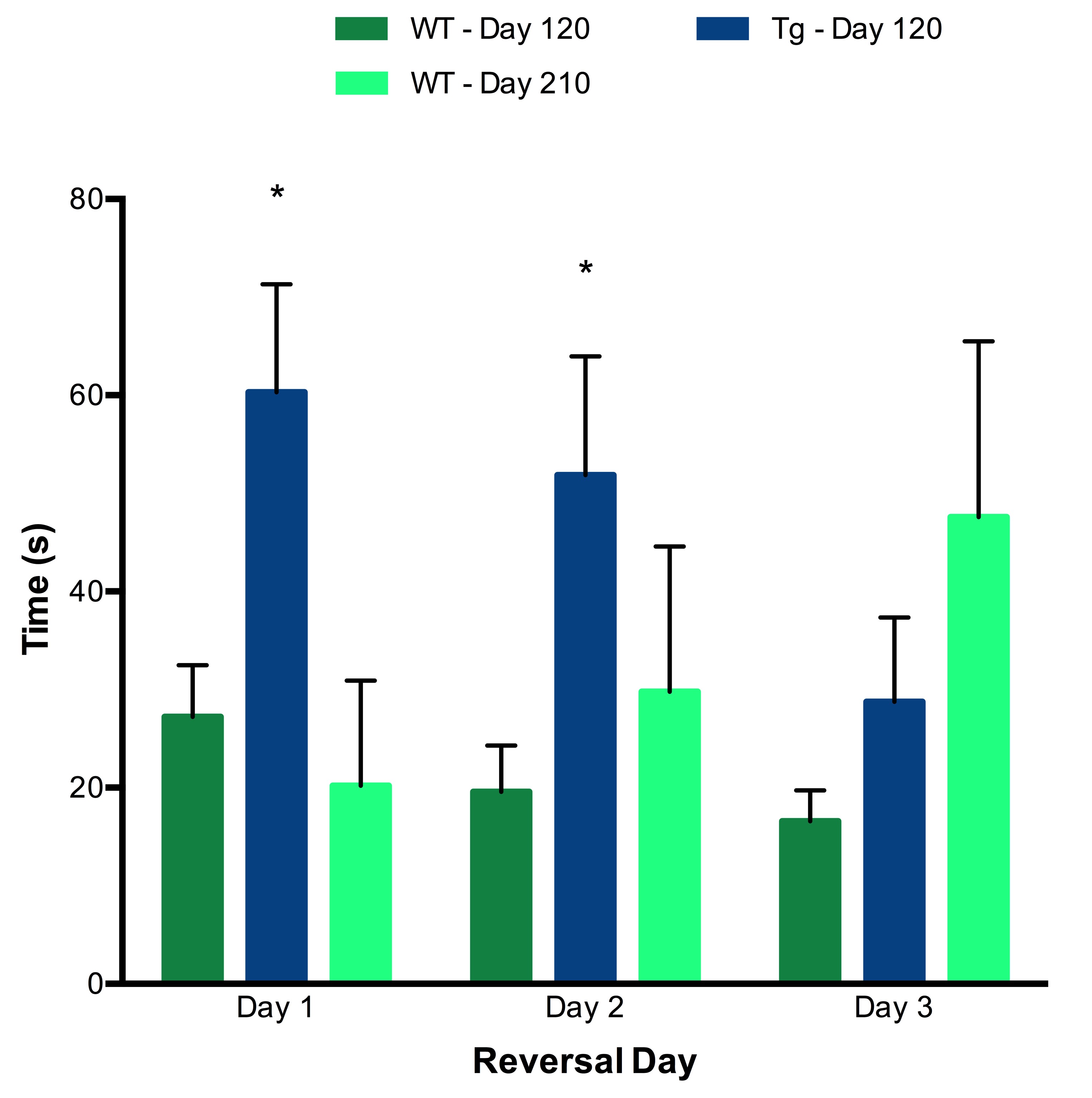
A Barnes maze consisting of 20 evenly spaced holes around a circular platform with a single escape hole was used to measure behaviour. Spatial cues served for orientation. There were four days of training (4 trials/day), one probe trial 72 hours after with the escape hole removed, and three days of reversal with the escape hole rotated (4 trials/day).5,6 For training and reversal, latency to target, path length and errors were recorded with Ethovision.9 For probe, time spent and holes searched per quadrant were recorded.
Solid-phase sandwich ELISAs from ThermoFisher were used to quantify plasma levels of the cytokines TNF-α (#KRC3011), IL-1β (#BMS630) and IL-6 (#BMS625). A colourimetric microplate reader was used for quantification.
Results
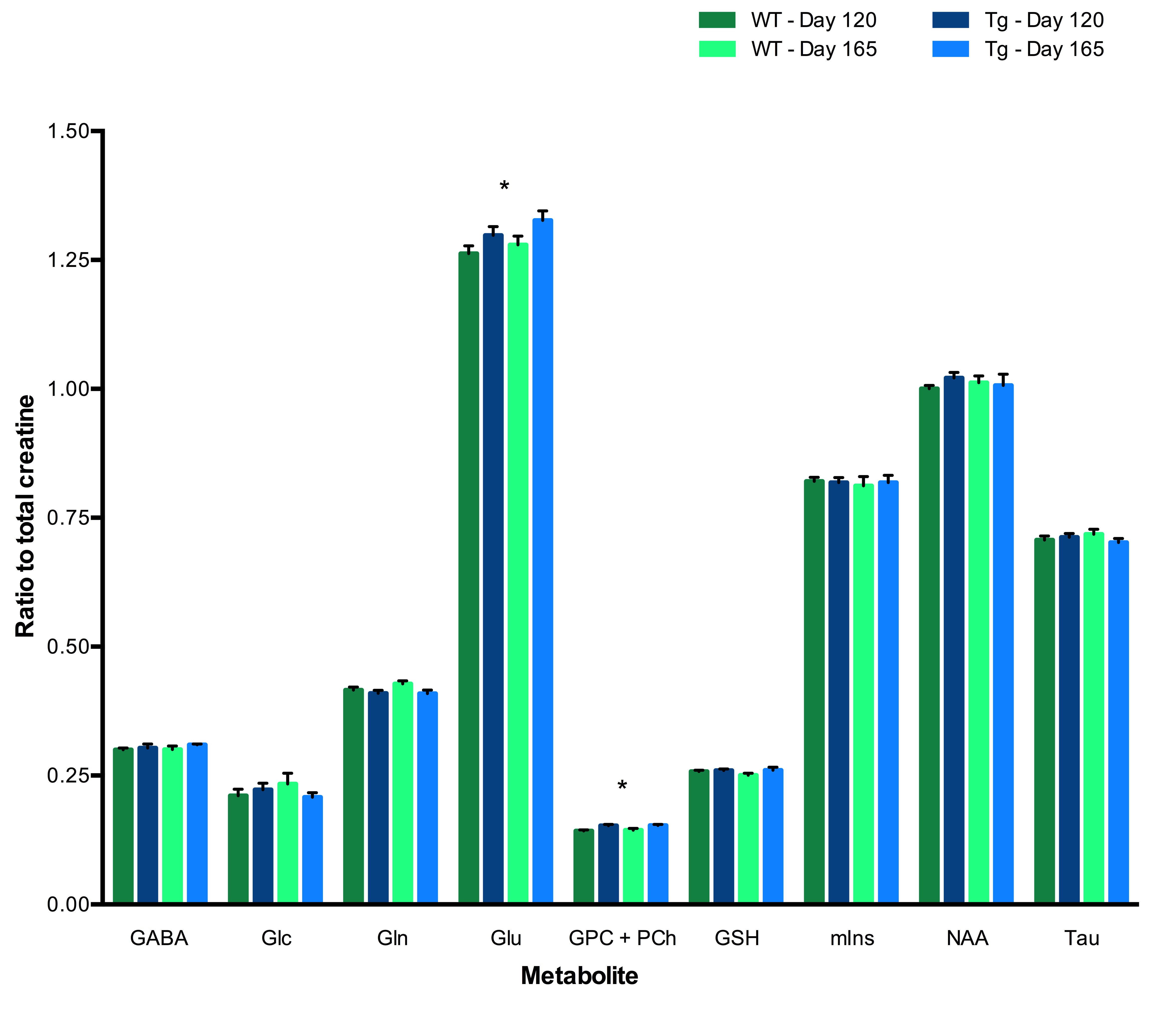
Tg rats showed significantly elevated levels of Glu and total choline (GPC+PCh) compared to wild-type (WT) rats at both Day 120 and Day 165. No effects of either genotype or time were observed for any other metabolites (Figure 2). Early behavioural results indicate that as early as 120 days, Tg rats perform worse on both the training trials (Figure 3) and reversal trials (Figure 4) of the Barnes maze compared to WT rats. Analysis of ELISA measures of systemic cytokine levels is currently in progress.Discussion
Early neurochemical alterations were observed in Tg rats relative to WT controls. Compared to later disease states, where reductions in Glu and N-acetylaspartate (NAA) have been observed and interpreted as relating to neuronal loss, here we observe a paradoxical increase in Glu in Tg rats.3,10 This may represent increased neuronal activity noted in prodromal AD, possibly triggered by elevated proinflammatory cytokines.2,3 Ongoing analysis of plasma cytokine levels will address this possible relationship. In addition, elevated levels of GPC + PCh, related to membrane turnover, may indicate impaired homeostasis or cellular proliferation. At this early stage, no change in mIns or NAA was observed.
Behavioural data suggest that Tg rats experience early cognitive decline. The concurrently observed neurotransmitter imbalance may underlie these behavioural alterations. Further analyses are in progress to characterize the early cytokine profile in this rodent model, in order to understand the relationships between cytokine activity, early neurochemical changes and cognitive dysfunction. This ongoing study represents a first step towards understanding the impact of abnormal systemic cytokine levels on brain chemistry and function in AD.
Conclusion
These preliminary findings highlight the potential for MRS to identify and monitor subtle changes in neurochemistry, and to relate these changes to other biochemical and behavioural alterations as a way to understand their collective role in the cascade of AD pathology. This multi-dimensional approach may provide a means to identify biomarkers for early diagnosis, possibly leading to improved treatment.Acknowledgements
This study was funded by CIHR and FRQS awarded to Dr. Jamie Near, in addition to a fellowship for graduate students from Healthy Brains for Healthy Lives awarded to Katrina Cruickshank.References
1. Blennow K, de Leon M, & Zetterberg H. Alzheimer’s disease. Lancet. 2006;368(9533):387-403.
2. Cavanagh C, Colby-Milley J, Bouvier D, et al. βCTF-correlated burst of hippocampal TNFα occurs at a very early, pre-plaque sage in the TgCRND8 mouse model of Alzheimer’s disease. J Alzheimer’s. 2013;36(2):233-238.
3. Cavanagh C, Chung Tse Y, Nguyen H, et al. Inhibiting tumor necrosis factor-α before amyloidosis prevents synaptic deficits in an Alzheimer’s disease model. Neurobiol Aging. 2016;47:41-49.
4. Marjanska M, Curran G, Wengenack T, et al. Monitoring disease progression in transgenic mouse models of Alzheimer’s disease with proton magnetic resonance spectroscopy. PNAS. 2005;102(33):11906-11910.
5. Cohen R, Rezai-Zadeh K, Weitz T, et al. A transgenic Alzheimer rat with plaques, tau pathology, behavioural impairment, oligomeric Aβ, and frank neuronal loss. J Neuro. 2013;33(15):6245-6256.
6. Barnes C. Memory deficits associated with senescence: Neurophysiological and behavioural study in the rat. J Comp Physiol. 1979;93(1):74-104.
7. Simpson R, Devenyi G, Jezzard P, et al. Advanced processing and simulation of MRS data using the FID appliance (FID-A) - An open source, MATLAB-based toolkit. Magn Reson Med. 2015;77(1):23-33.
8. Provencher S. Automatic quantitation of localized in vivo 1H spectra with LCModel. NMR Biomed. 2001;14:260-264.
9. Noldus L, Spink A, & Tegelenbosch R. A versatile video tracking system for automation of behavioural experiments. Behav Res Methods Instrum Comput . 2001;33(3):398-414.
10. von Kienlin M, Künnecke B, Metzger F, et al. Altered metabolic profile in the frontal cortex of PS2APP transgenic mice, monitored throughout their lifespan. Neurobiol Dis. 2005;18(1):32-39.
Figures