2255
MR compatible cell perfusion system allows investigating metabolism and invasion of hepatocellular carcinoma cells during p53 re-activation induced senescence1Division of Cancer Imaging Research, The Russell H Morgan Department of Radiology and Radiological Science, The Johns Hopkins University, School of Medicine, Baltimore, MD, United States, 2Werner Siemens Imaging Center, Department of Preclinical Imaging and Radiopharmacy, Eberhard Karls University Tübingen, Tübingen, Germany, 3Department of Internal Medicine VIII, Eberhard Karls University Tübingen, Tübingen, Germany, 4Sidney Kimmel Comprehensive Cancer Center, The Johns Hopkins University, School of Medicine, Baltimore, MD, United States, 5Department of Radiation Oncology and Molecular Radiation Sciences, The Johns Hopkins University, School of Medicine, Baltimore, MD, United States
Synopsis
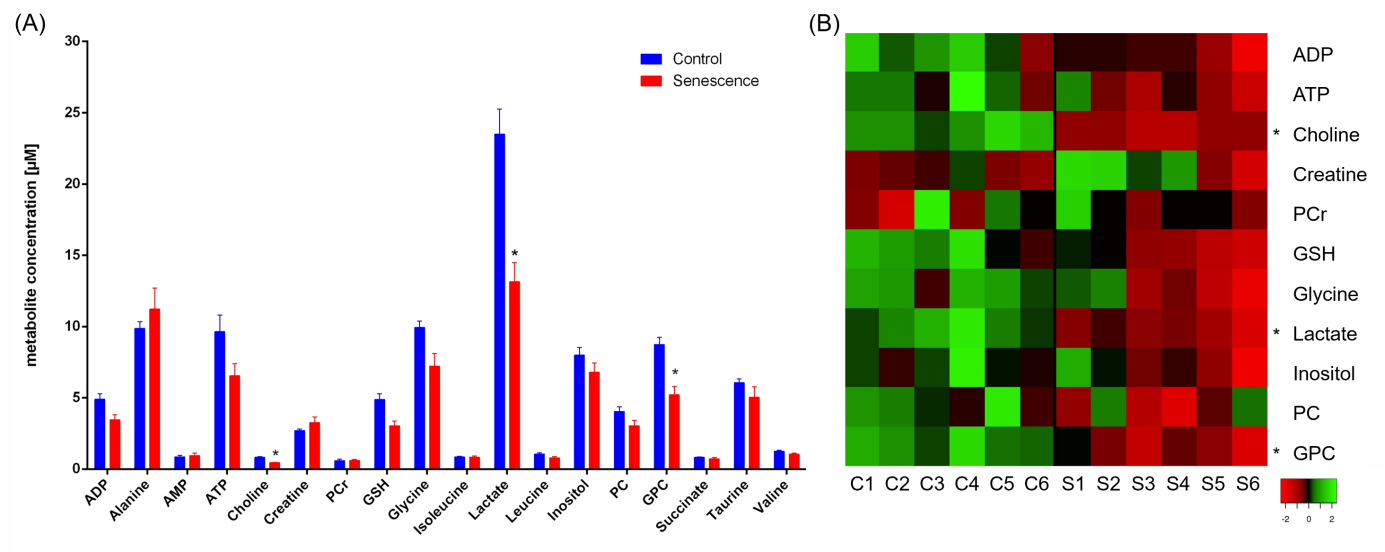
Here we investigated the metabolic characteristics and the effect of the so called senescence associated secretory phenotype (SASP) on extracellular matrix (ECM) degradation of p53 re-activation induced senescence, using a murine liver carcinoma cell model, where p53 is silenced in the presence of doxycycline hyclate. Senescence is induced within three days after doxycycline hyclate withdrawal and subsequent p53 re-activation. Our data revealed alterations of metabolites in p53-reactivation induced senescent H-Ras cells compared to control cells, including creatine, phosphocreatine and glycerophosphocholine indicating differences in energy and phospholipid metabolism. Senescent H-Ras cells tended to degrade the ECM more than control cells.
Introduction
Cellular senescence is characterized by a stable cell-cycle arrest. In literature replicative senescence, DNA damage-induced senescence and oncogene-induced senescence have been described. Induction of senescence in tumor cells could be either beneficial or detrimental in tumors. A characteristic feature of senescent cells is the so-called senescence associated secretory phenotype (SASP) consisting of cytokines, proteases and different growth factors. Therefore, understanding the metabolic characteristics of senescent cells and the effect of the SASP on invasion and extracellular matrix (ECM) degradation, can reveal insights into molecular processes and potentially lead to the development of novel treatments.Methods
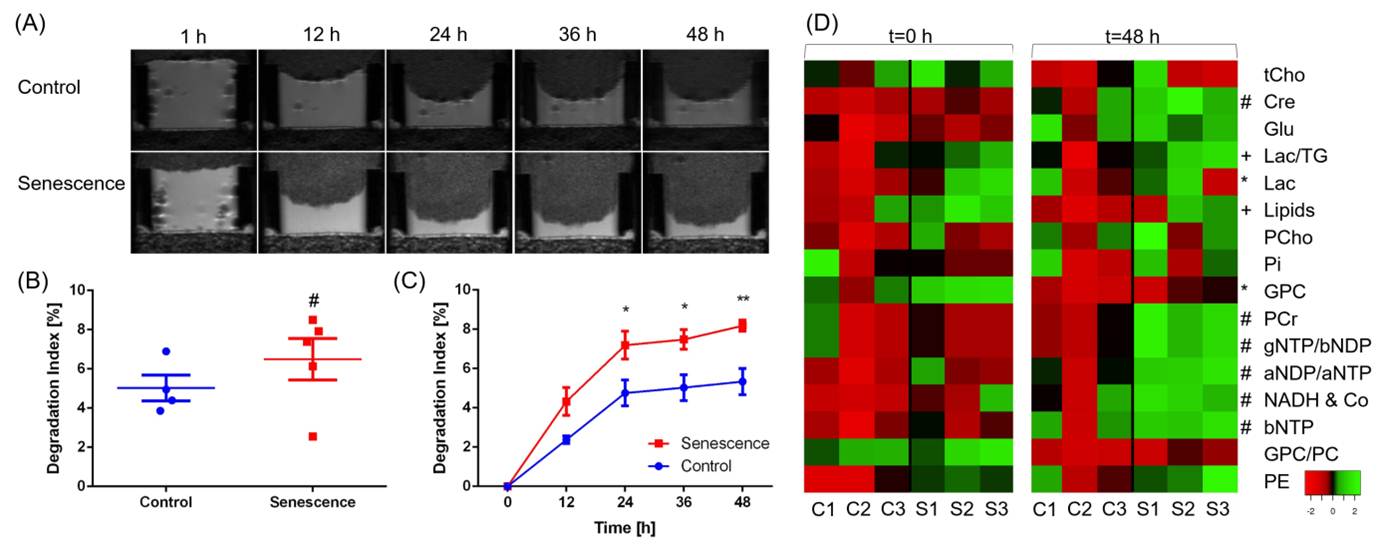
Here we investigated the metabolic characteristics of p53 re-activation induced senescence, using a murine liver carcinoma cell model (shp53;HRasV12, short H-Ras), where p53 is silenced in the presence of doxycycline hyclate. Senescence is induced within three days after doxycycline hyclate withdrawal and subsequent p53 re-activation. Ex-vivo analysis of control and senescent (three days after senescence induction) hepatocarcinoma H-Ras cells were performed on a high-resolution 1H NMR spectrometer (600 MHz Bruker Avance III HD) equipped with a triple resonance 5 mm TXI probe. Cell extracts were prepared using a 2-phase MeOH:MTBE solvent system. Spectra of the polar phase (six biological replicates from senescent and control cells) were analyzed and processed using TopSpin 3.5, metabolites annotated and quantified using ChenomX NMR Suite 8.3. Statistical significance was evaluated using the Student’s t-test. For metabolic and degradative investigations of H-Ras cells during senescence, an MR compatible cell perfusion was used. H-Ras cells were cultured on micro-carrier beads for 4 days, loaded into a cell perfusion system and monitored for 48 h, as previously described [1]. MR data were acquired on a 9.4 T MR spectrometer (Bruker, Billerica, MA). T1-weighted 1H MR imaging was performed to evaluate the sample preparation, to visualize the geometry of the ECM gel, and to detect changes in its integrity due to invasion and degradation. One-dimensional (1D) 1H MR profiles of intracellular water were acquired along the length (z-axis) of the sample by diffusion-weighted (DW) MRI. Intracellular metabolite levels were also derived from unlocalized DW 1H MR spectra. The first contact of cancer cells with the ECM gel during the loading of the sample was defined as the zero time point. All MR data were processed using XsOs MR Software. Values are presented as Mean ± SEM of at least three independent measurements.Results
Various metabolites were altered in p53 re-activation induced senescent cells compared to non-senescent cells (Figure 1). Control cells were high in lactate, glycine and choline derivate peaks, whereas senescent cells were high in alanine and creatine peaks. This indicates a general reduced metabolism in senescence. A principal component analysis (PCA) of all processed spectra showed a clear separation between the two groups. Some of the identified compounds were characterized by strong singlet peaks in the 1H NMR spectra that may have the potential to be detected by clinical MRS or to serve as targets for new tracer development. Using our MR compatible cell perfusion system, we found that p53-reactivation induced senescent hepatocellular carcinoma cells show a higher degradation index compared to control cells indicating an increase in ECM degradation (Figure 2). We also detected changes in the metabolome from both senescent and control cells and how they evolve with time.Discussion
Our data revealed alterations of metabolites in p53-reactivation induced senescent H-Ras cells compared to control cells, including creatine, phosphocreatine and glycerophosphocholine indicating differences in energy and phospholipid metabolism. Furthermore senescent H-Ras cells tended to degrade the ECM more than control cells.Conclusion
Metabolite profiles of senescent and non-senescent H-Ras cells can be characterized using ex-vivo high-resolution 1H NMR spectroscopy of cell extracts. The use of a MR compatible cell perfusion system furthermore allows investigations of metabolic alterations together with dynamic quantification of ECM degradation during senescence of intact perfused cells. The study provides insights into the metabolic processes underlying senescence in cancer.Acknowledgements
PK was supported by a grant from the German Academic Exchange Service (DAAD PPP USA 2018, Project-ID 57387312). JPT was supported by Alonso Martin Escudero foundation. Studies supported by NIH R35 CA209960.References
1. Ackerstaff E, Artemov D, Gillies RJ, Bhujwalla ZM. Hypoxia and the presence of human vascular endothelial cells affect prostate cancer cell invasion and metabolism. Neoplasia. 2007;9(12):1138-1151.Figures