1283
Simulated effect of diffusion time and skeletal muscle fiber size on the diffusion tensorDavid Barnes Berry1, Erin Englund1, Vitaly Galinsky1, Chamindra Konersman1, Shaochen Chen1, Samuel Ward1, and Lawrence Frank1
1University of California, San Diego, La Jolla, CA, United States
Synopsis
The sensitivity of diffusion tensor imaging (DTI) to muscle fiber size is dependent upon diffusion time. However, there is no consensus on how to interpret data acquired at different diffusion times. In this study we simulated the relationship between muscle fiber size, diffusion time, and the resulting diffusion tensor in models with simplified and histology informed muscle fiber geometry, using a stimulated echo DTI pulse sequence. Maximum contrast between physiologically relevant fiber sizes was found at 130ms for diffusivity, and 170ms for fractional anisotropy measurements. This data may better inform pulse sequence parameter selection when performing DTI experiments in vivo.
Introduction
Diffusion tensor imaging (DTI) has been used to assess restricted diffusion in skeletal muscle and is thought to track microstructural, and thereby functional changes. Stimulated echo DTI can be used to probe muscle microstructure at long diffusion times, which enhances diffusion contrast.1 As sensitivity to muscle fiber size is dependent on the diffusion time chosen, it is important to optimize diffusion pulse sequence parameters in order to maximize diffusion contrast of physiologically relevant muscle fiber sizes. However, many studies seem to arbitrarily select diffusion time since the relationship between diffusion time, muscle fiber size, and the diffusion tensor is unknown and difficult to precisely test in vivo. In this study, we used an in silico approach to explore the relationship between muscle fiber diameter and the diffusion tensor at a range of diffusion times, in models with simplified, and histology-informed geometry. Additionally, we investigated at what diffusion time we observed the greatest contrast in diffusion measurements of physiologically relevant fiber sizes.Methods
The MRI simulation tool DifSim was used to model DTI experiments to study the relationship between muscle fiber size, and the diffusion tensor at different diffusion times. DifSim employs MCell, a Monte Carlo simulator for cellular microphysiology, to simulate the diffusion of particles, and tracks particle location, signal amplitude and phase, with respect to a specified MR pulse sequence, within a user-defined, arbitrarily complex model.2 A stimulated echo DTI pulse sequence was simulated, with the following sequence parameters: TE=21.76ms; b=500s/mm2; voxel size=200×200×200μm3; directions=15; δ=2ms; Δ=20ms, 30ms, 40ms, 50ms, 90ms, 130ms, 170ms, 250ms, 325ms, 400ms, 500ms, and 750ms. The diffusion tensor was calculated using AFNI.3 Muscle fibers with ideal geometry were approximated as tightly packed hexagons, surrounded by ECM (Figure 1).4 Muscle fiber size was varied by systematically changing the diameter of the hexagons over a physiologically informed range (10-100μm). Models with histology informed geometry were derived from control, botox injected, surgically denervated, and surgically tenotomized rat tibialis anterior muscles 30 days after injury (Figure 2). Model inputs for diffusion coefficients and relaxation times are reported in Table 1.To determine at what diffusion time the largest contrast between physiologically relevant fiber sizes was observed, we calculated the difference between 40μmand 60μmdiameter simplified models, and the largest and smallest diameter histology informed models at all diffusion times. These fiber sizes were selected as it is roughly the average size of a human muscle fiber, and the range at which atrophic or hypertrophic changes occur.5Results
As diffusion time increased, fractional anisotropy increased, while mean and radial diffusivity decreased for all models (Figure 3A-C, 4A-C). In the simplified muscle models, the largest difference in mean and radial diffusivity between 40μm and 60μm diameter fibers was found at Δ=130ms (19%), while the largest difference in fractional anisotropy was found at Δ=170ms (43%; Figure 3D-F). Similarly, the greatest difference between the largest (control – 63.8μm) and smallest (botox – 34.0μm) histology informed models was found at Δ=130ms for mean and radial diffusivity (24%), and Δ=170ms for fractional anisotropy (70%; Figure 4D-F).Discussion
In this study, we have described, with a series of highly controlled simulations, the relationship between muscle fiber size, diffusion time, and the diffusion tensor. We identified a range of diffusion times (130-170ms) at which the largest dynamic range in diffusion measurements was found for physiologically relevant fiber sizes and geometry. Overall, differences between muscle fiber sizes were much greater for fractional anisotropy, than diffusivity measurements, suggesting that fractional anisotropy may be more sensitive to muscle microstructure than diffusivity measurements. Muscle fiber size is correlated with force production in muscle, and atrophic changes are coincident with injury and myopathic disease progression. Therefore, the main goal of DTI in skeletal muscle is to precisely understand the relationship between the diffusion tensor and underlying microstructure. While sampling multiple diffusion times in an in vivo experiment may better describe underlying muscle structure,6 this may not be feasible in all experiments due to inherent long scan times associated with DTI. Therefore, if only a single diffusion time can be assessed, then the selection of diffusion time should be informed by the time at which the greatest contrast may be present.Conclusions
These findings describe the relationship between fiber size and the diffusion tensor as a function of diffusion time. We demonstrate that at a diffusion time of Δ=130ms-170ms, the dynamic range of diffusion measurements is maximized at physiologically relevant fiber sizes (40μm-60μm), which may better inform pulse sequence parameter selection when performing DTI experiments of skeletal muscle in vivo.Acknowledgements
The authors would like to thank Keaton Blazer for his help in generating the histology-informed computer models. This work was supported by R01 AR070830.References
- Sigmund EE, Sui D, Ukpebor O, et al. Stimulated echo diffusion tensor imaging and SPAIR T2-weighted imaging in chronic exertional compartment syndrome of the lower leg muscles. JMRI. 2013;38:1073–1082.
- Balls GT and Frank LR. A simulation environment for diffusion weighted MR experiments in complex media. MRM, 2009;62(3):771-778.
- Cox RW. AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Comp Biomed Res. 1996;29:162–173.
- Berry DB, Regner B, Galinsky V, et al. Relationships between tissue microstructure and the diffusion tensor in simulated skeletal muscle. MRM. 2018;80:317-329.
- Lieber RL. Skeletal muscle structure, function, and plasticity. Philadelphia, PA, USA: Lippencott Williams & Wilkins; 2002.
- Winters KV, Reynaud O, Novikov DS, Fieremans, E. Quantifying myofiber integrity using diffusion MRI and random permeable barrier modeling in skeletal muscle growth and Duchenne muscular dystrophy model in mice. MRM. 2018;80:2094-2108.
- Fan RH and Does M.D. Compartmental Relaxation and DTI Measurements In Vivo in λ-Carrageenan Induced Edema in Rat Skeletal Muscle. NMR Biomed, 2008;21(6):566-573.
- Crémillieux Y, Ding S, and Dunn JF, High‐resolution in vivo measurements of transverse relaxation times in rats at 7 Tesla. MRM, 1998;39(2):285-290.
- Diakova, G, Korb JP, and Bryant RG, The magnetic field dependence of water T1 in tissues. MRM, 2012;68(1):272-277.
- Gefen A, Cornelissen LH, Gawlitta D, et al. The free diffusion of macromolecules in tissue-engineered skeletal muscle subjected to large compression strains. J Biomech, 2008;41(4):845-853.
- Moll W. The diffusion coefficient of myoglobin in muscle homogenate. Pflügers Arch Gesamte Physiol Menschen Tiere. 1968;299(3):247-251.
- Papadopoulos S, Jürgens KD, and Gros G. Protein diffusion in living skeletal muscle fibers: dependence on protein size, fiber type, and contraction. Biophys J. 2000;79(4):2084-2094.
- Gilbert DL, Okano T, Miyata T, Kim SW. Macromolecular diffusion through collagen membranes. Int J Pharm. 1988;47(1):79-88.
- Ramanujan S, Pluen A, McKee TD, et al. Diffusion and convection in collagen gels: implications for transport in the tumor interstitium. Biophys J. 2002;83(3):1650-1660.
Figures

Figure 1. Schematic depicting simplified hexagonal models of skeletal muscle (top row; intracellular) and the ECM (middle row; extracellular). Changes in fiber size was simulated by changing the diameter of the muscle fibers.
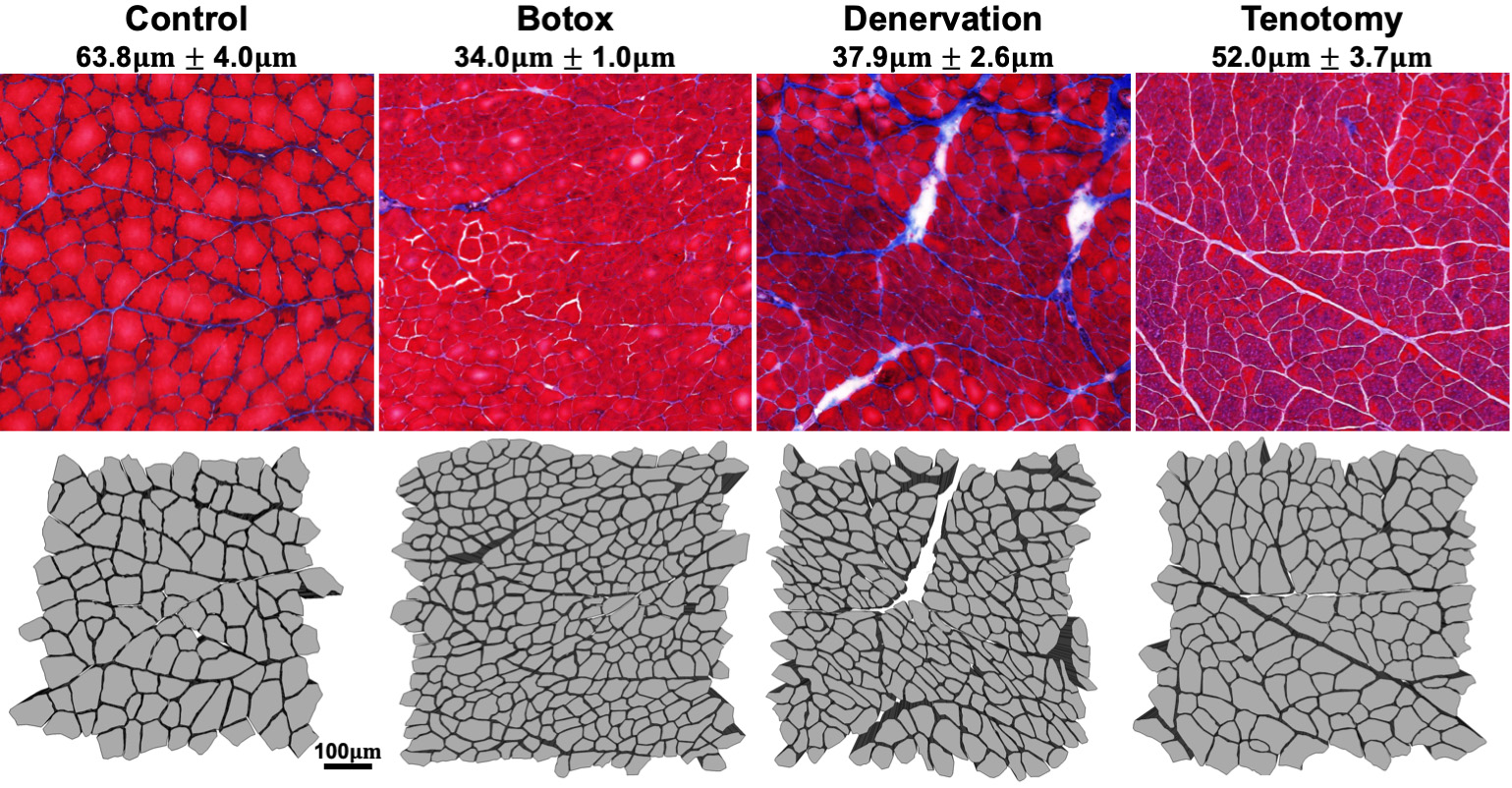
Figure 2. Schematic depicting histology-informed models of skeletal muscle. Fibers were manually traced from histology images (top row). Botox, denervation, and tenotomy models were taken 30 days post injury. Mean +/- standard deviation of fiber size is reported for each injury model.
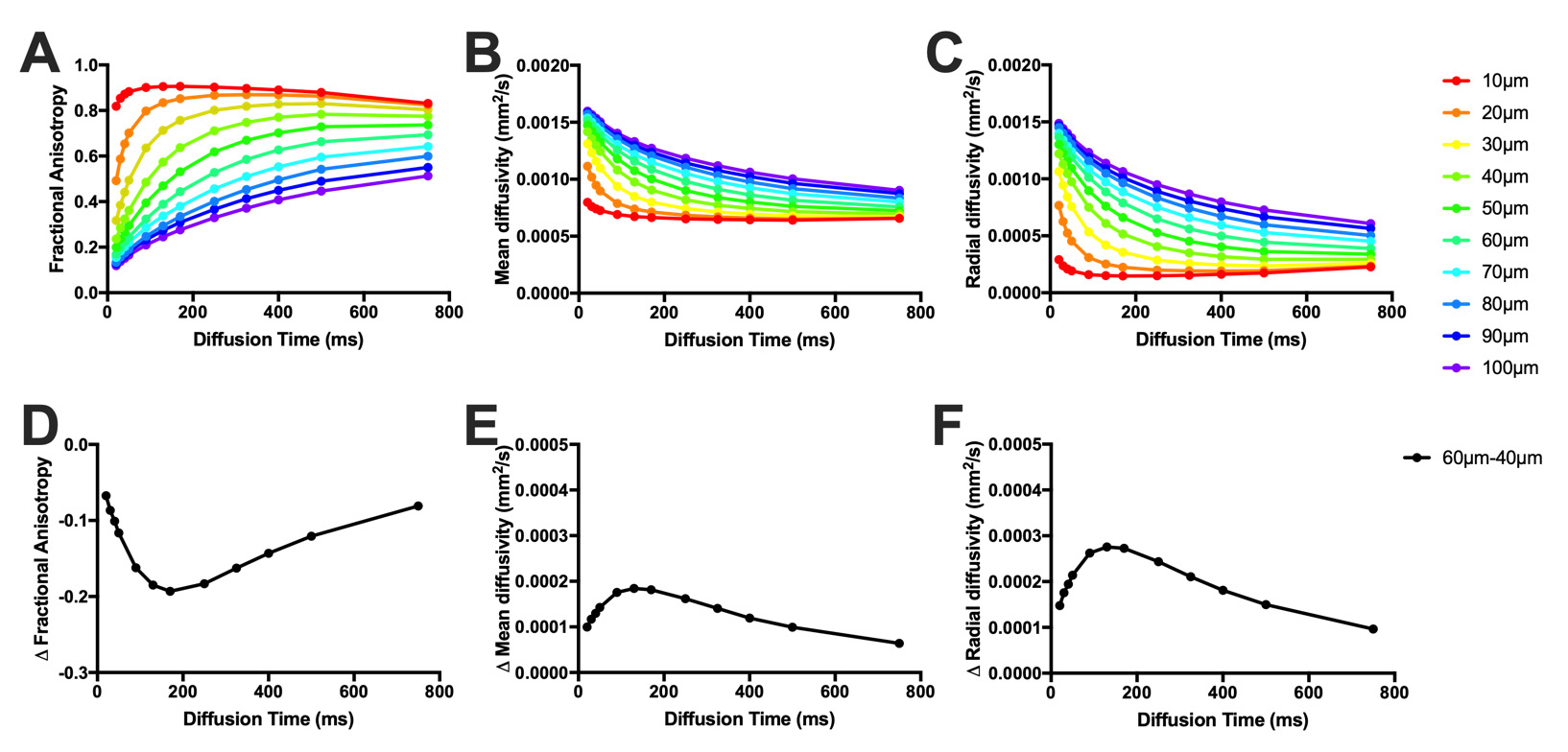
Figure 3. A) Fractional anisotropy, B) mean diffusivity, and C) radial diffusivity of simplified muscle models with diameter 10μm-100μm at diffusion times ranging from 20ms-750ms. The difference in D) fractional anisotropy, E) mean diffusivity, and F) radial diffusivity between 40μm-60μm diameter simplified models was calculated in order to determine the dynamic range of measurements derived from the diffusion tensor as a function of diffusion time at common muscle fibers sizes. The greatest contrast between 40μm-60μm fibers was found at Δ=130ms for diffusivity measurements, and Δ=170ms for fractional anisotropy.
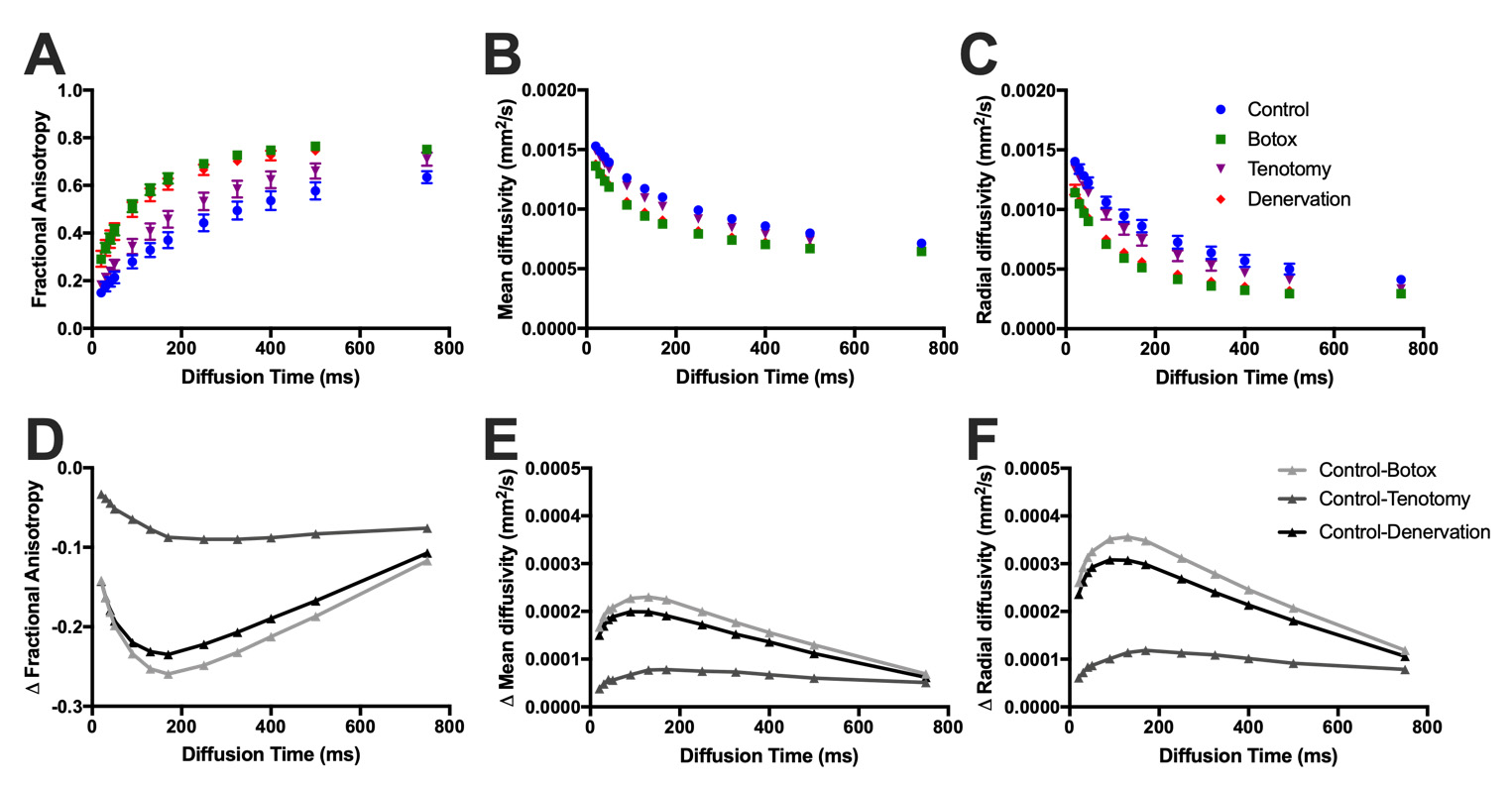
Figure 4. A) Fractional anisotropy, B) mean diffusivity, and C) radial diffusivity of histology-informed muscle models from control, botox, tenotomy, and denervated animal models at various diffusion times. Difference in D) fractional anisotropy, E) mean diffusivity, and F) radial diffusivity between the control and injury models was calculated to determine the dynamic range of measurements as a function of diffusion time in histology-informed geometry models. The greatest contrast was found at 130ms for diffusivity measurements, and 170ms for fractional anisotropy between control and small muscle fiber models (botox, denervation), but was 325ms and 170ms respectively for similarly sized models (tenotomy).

Table 1. Relaxation times and diffusion coefficients of intra- and extracellular components of the muscle models taken from the literature.