1051
Triple network activity regulation mediated by the insular cortex in the mouse brain.1Singapore Bioimaging Consortium, Agency for Science, Technology and Research, Singapore, Singapore, 2Faculty of Biology, Medicine and Health, The University of Manchester, Manchester, United Kingdom, 3University College London, London, United Kingdom
Synopsis
The triple-network model is a contemporary theoretical framework derived from empirical neuroimaging data to explain a wide range of observations stemming from multiple psychopathologies. Central to the model are interactions between the salience, default-mode, and central executive networks. The insula area is a central node of the salience network. Using acute photostimulation, we report evidence to support the existence of a triple-network system within the mouse brain. Further, by using sustained optogenetic neuromodulation, we show that inhibition of the insular area acts on homotopic functional connectivity but fails to affect wider interactions within triple-network system in a resting-state setting.
Introduction
Mental disorders are increasingly perceived as distributed neuronal network disorders1. Among the networks affected, the salience network (SN) stands out as a vulnerable hub across several psychopathologies2. This network includes intrinsically connected regions at the interface of sensory, emotional and cognitive integration3,4. Its contribution to the healthy and diseased brain is at the center of the triple-network model5,6, a framework regrouping empirical findings from a number of psychopathologies into a unified model. In this model, the elements of the SN carry out salience mapping of external stimuli and exert dynamic switching between self-referential activity within the default-mode network (DMN) and externally-oriented activity associated with the central executive network (CEN). In this study, we aimed to determine whether: (i) the mouse brain presents a functional organization akin to the humain SN; (ii) acute photoactivation of the insula with optogenetics activates elements of the rodent DMN and CEN; and (iii) sustained neuromodulation of the insula may be used to manipulate these interactions.Method
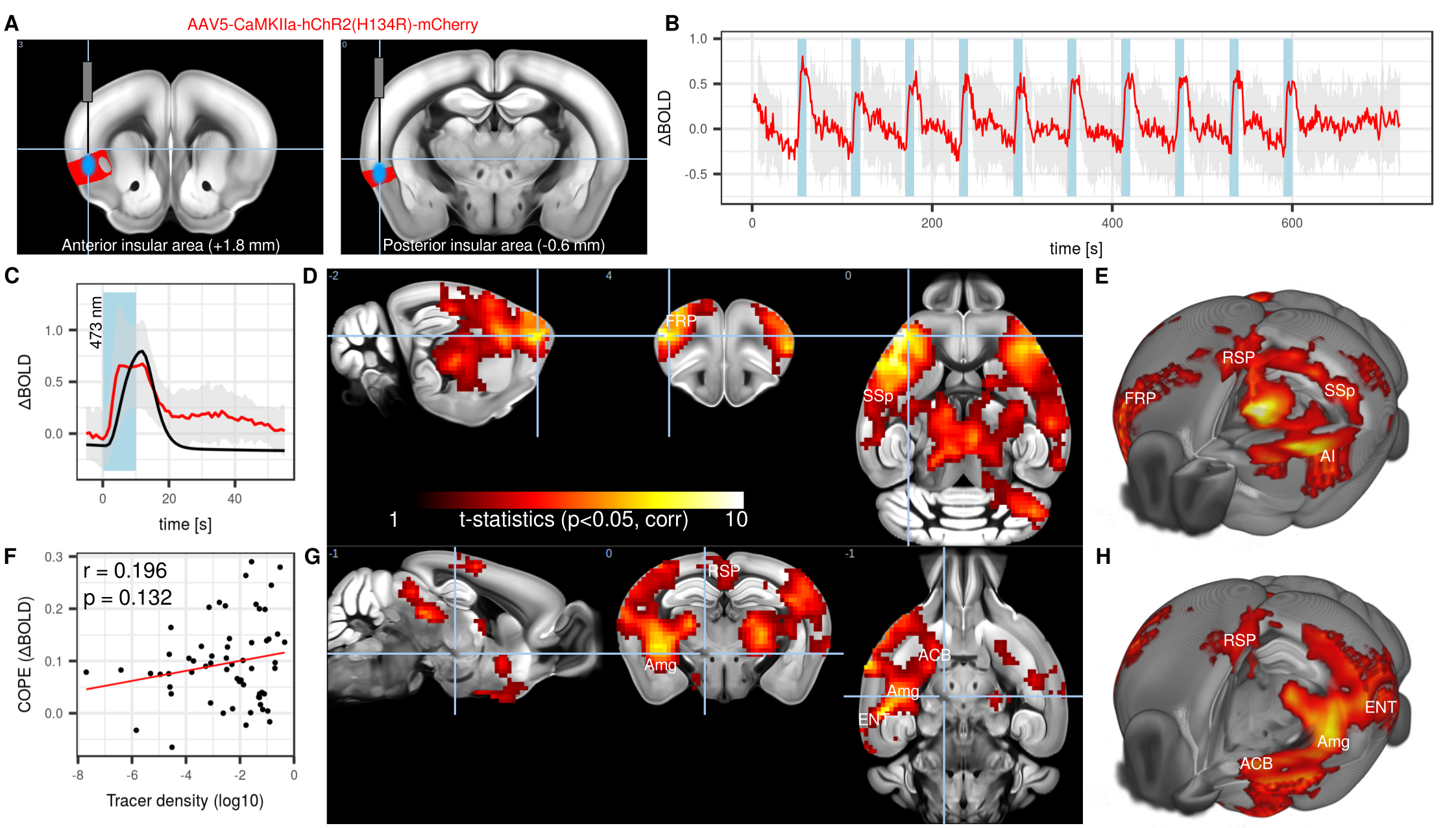
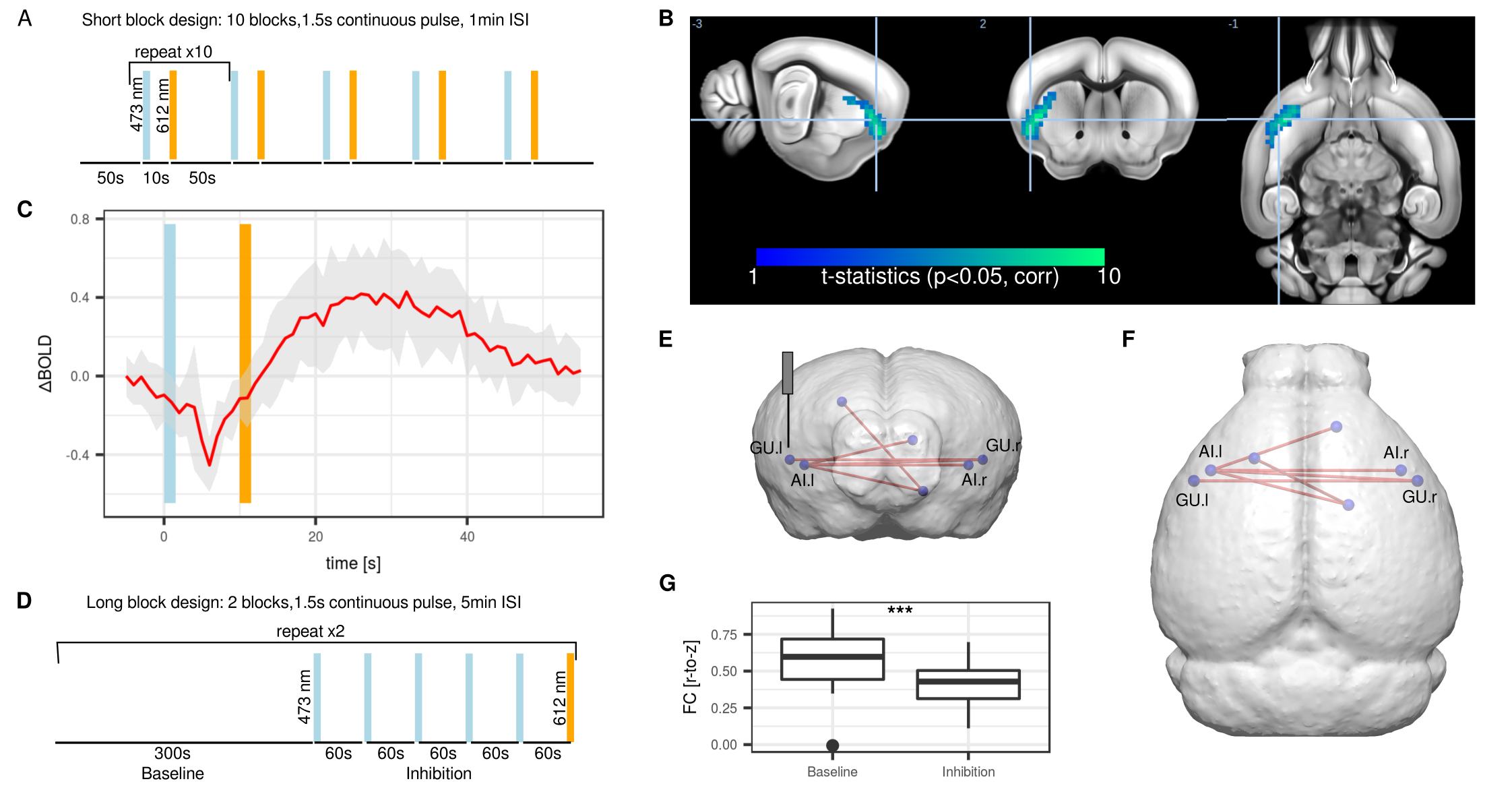
Nine C57Bl/6 mice were stereotactically injected with Channelrhodopsin 2 (ChR2, AAV5-CamKIIa-hChR2) in the left agranular insular area (AI), either in the frontal (+1.8 mm relative to Bregma) or posterior ( -0.6 mm) area (Figure 1A). A further 10 mice were injected with inhibitory stabilized step function opsin (SSFO, AAV5-hSyn-SwiCa++). An optical implant was inserted and secured with dental cement. Anesthetized mice underwent functional magnetic resonance imaging (fMRI) at 11.75T using a 10 mm surface receiver coil. Gradient-echo echo planar images were acquired with the following parameters TR = 1000 ms, TE = 11.7 ms, FA = 60°, matrix 60 x 30, FOV 17 x 9 mm, slice thickness 0.45 mm. Animals were imaged under isoflurane 0.5%, medetomidine 0.1 mg/kg/h, and muscle relaxation7. ChR2 was photostimulated with 10 s blocks at 473 nm, 20 Hz, 10 ms pulse width (Figure 1B). SSFO was photostimulated with 1.5 s blocks of continuous light at 473 nm for inhibition and 612 nm for reversal (Figure 2AD). Images were corrected for motion and smoothed (0.45 mm2). In stimulus-evoked condition, images were high-pass filtered (0.1 Hz) and a general linear model (GLM) analysis with gamma function (Figure 1C) was applied. In resting-state condition, vascular + ventricle signal regression together with band-pass filter (0.01-0.25 Hz) was applied followed by network analysis using predefined regions-of-interest.Results
Photostimulation of ChR2 lead to a 1% BOLD signal increase within the AI (Figure 1B). Group-level analysis of GLM of mice injected in the frontal AI revealed significant clusters in the frontal pole area (FRP), an area associated with the mouse CEN8, as well as retrosplenial area (RSP), an area associated with DMN (Figure 1DE). Comparatively, photostimulation of the posterior AI elicited response in the RSP and entorhinal area (ENT), other elements of the DMN, as well as amygdala (Amg) and nucleus accumbens (ACB), both elements of the SN (Figure 1GH). A low correlation (r = 0.196) between functional maps from the frontal AI with a corresponding viral tracer map derived from Allen Institute for Brain Science suggested the presence of poly-synaptic interactions (Figure 1F). We concluded that acute photostimulation of the AI leads to evoked response within the direct projection field of the AI and also within a greater ensemble of regions associated with the SN, DMN, and CEN, consistent with the concept of a rodent triple-network system. To achieve sustained neuromodulation, inhibitory SSFO was used instead. A short block protocol (Figure 2A) lead to a 0.4% BOLD decrease (Figure 2B). The response was confined to the area under the optical implant (Figure 2C). A sustained inhibition protocol comprising of 4 interleaved blocks of 5 min baseline and 5 min inhibition was used in a resting-state setting (Figure 2D). Comparisons between baseline and SSFO-mediated inhibition indicated reduction in homotopic functional connectivity (FC) in AI and gustatory area (GU) (Figure 2EF). Contralateral FC was reduced by 45% during inhibition (Figure 2G). Interestingly, non-homotopic FC was unaffected by local inhibition of the left AI, including regions that were highlighted following ChR2 photostimulation.Conclusion
In summary, we demonstrate evidence for a triple-network model in the mouse. These interactions are hypothesized to hold particular relevance in animal models of psychopathologies, comparable to reports in human studies. The mouse offers a system to investigate these interactions in detail using cellular-specific neuromodulation tools such as SSFO.Acknowledgements
This work was supported by the Singapore Bioimaging Consortium (SBIC), A*STAR, Singapore.References
1. Buckholtz, J. W. & Meyer-Lindenberg, A. Psychopathology and the human connectome: toward a transdiagnostic model of risk for mental illness. Neuron 74, 990–1004 (2012).
2. McTeague, L. M. et al. Identification of common neural circuit disruptions in cognitive control across psychiatric disorders. Am. J. Psychiatry 174, 676–685 (2017).
3. Manoliu, A. et al. Insular dysfunction within the salience network is associated with severity of symptoms and aberrant inter-network connectivity in major depressive disorder. Front. Hum. Neurosci. 7, 930 (2013).
4. Craig, A. D. B. How do you feel--now? The anterior insula and human awareness. Nat. Rev. Neurosci. 10, 59–70 (2009).
5. Menon, V. & Uddin, L. Q. Saliency, switching, attention and control: a network model of insula function. Brain Struct. Funct. 214, 655–667 (2010).
6. Menon, V. Large-scale brain networks and psychopathology: a unifying triple network model. Trends Cogn Sci (Regul Ed) 15, 483–506 (2011).
7. Grandjean, J., Schroeter, A., Batata, I. & Rudin, M. Optimization of anesthesia protocol for resting-state fMRI in mice based on differential effects of anesthetics on functional connectivity patterns. Neuroimage 102 Pt 2, 838–847 (2014).8. Gozzi, A. & Schwarz, A. J. Large-scale functional connectivity networks in the rodent brain. Neuroimage 127, 496–509 (2016).
Figures