1049
Submillimeter whole brain diffusion MRI at 7 Tesla using simultaneous multislice imaging1Center for Magnetic Resonance Research, Radiology, Medical School of the University of Minnesota, Minneapolis, MN, United States
Synopsis
Acquisition of whole-brain diffusion MRI at submillimeter resolutions has proven to be highly beneficial for probing the brain’s structure and connectivity. Here, we investigate how simultaneous multislice (SMS) imaging, a single-shot acquisition that has great success in large-scale cohort studies such as the Human Connectome Projects (HCP), can be used to achieve submillimeter whole-brain diffusion at 7 Tesla. We acquired SMS diffusion at 0.7-mm isotropic resolution using a standard body gradient. Our results show that these submillimeter data can be used to generate a whole-brain tractogram and a connectome comparable to those of the 1.05-mm HCP-style acquisition using denoising approaches.
Introduction
Acquisition of whole-brain diffusion MRI at submillimeter resolutions has proven highly beneficial for probing the brain’s structure and connectivity1.
Previous studies targeting in-vivo submillimeter diffusion have opted for 3D acquisitions (e.g., multi-slab2 or gSLIDER3) for improved SNR efficiency. However, these methods are intrinsically multi-shot and therefore prone to motion-induced shot-to-shot phase errors.
Here, we investigate how simultaneous multislice (SMS) imaging4,5, a pseudo-3D single-shot acquisition that has great success in large-scale cohort studies such as the Human Connectome Projects (HCP)6,7, can be used to achieve submillimeter whole-brain diffusion at 7 Tesla (7T).
We acquired SMS diffusion at 0.7-mm isotropic resolution using a standard body gradient and show that these submillimeter data can be used to generate a whole-brain tractogram and a structure connectome comparable to those of the 1.05-mm HCP-style acquisition8.
Methods
We collected human brain images on a Siemens 7T MR scanner, equipped with a standard body gradient (70 mT/m maximum strength and 200 T/m/s maximum slew rate) and 32-channel receive (32Rx) capability.
The system can be operated in a parallel-transmission (pTx) mode enabling up to 16 transmit channels.
A healthy subject who signed a consent form approved by the local IRB was scanned using the commercially-available Nova 8-channel transmit 32Rx coil and in the pTx “protected” mode to ensure RF safety.
Data acquisition:
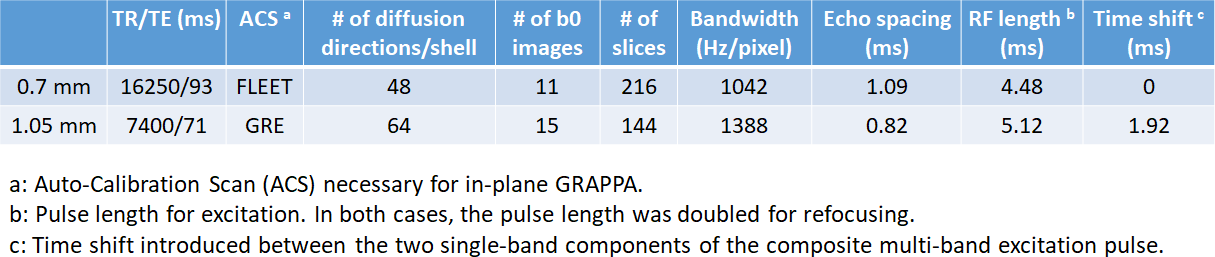
We acquired whole-brain diffusion at 0.7-mm and 1.05-mm8,9 isotropic resolutions (the latter serving as a reference). The two imaging protocols are compared in Fig. 1.
Briefly, both acquisitions sampled the q-space in two shells with uniform angular coverage10 and utilized slab-wise pTx multi-band pulses for improved RF uniformity across the brain8.
While the 1.05-mm acquisition continued to use the GRE auto-calibration scans (ACS) for in-plane GRAPPA as in the HCP9, the 0.7-mm acquisition opted for the FLEET ACS11 which was found to minimize residual aliasing in final reconstructed images.
For both acquisitions, diffusion images of individual receive channels were reconstructed using the split slice-GRAPPA algorithm12 to minimize inter-slice signal leakage and were combined to form the final image via the SENSE1 method13 to reduce the noise floor.
Data processing and analysis:
Both datasets were processed as follows:
1) denoising with the MP-PCA method14;
2) preprocessing using the HCP minimal pipelines15;
3) estimation of fiber orientation distribution functions (fODFs) using a multi-shell multi-tissue constrained spherical deconvolution approach16,17 coupled with unsupervised multi-tissue response function estimation18 and followed by multi-tissue informed intensity normalization;
4) reconstruction of a whole-brain tractogram with improved biological accuracy by combining a probabilistic streamlines algorithm19 with the anatomically-constrained tractography framework20 and spherical-deconvolution-informed filtering of tractography (SIFT2)21;
5) generation of a structural connectome22,23 for which the edges were calculated based on the whole-brain tractogram whereas the nodes (or gray matter parcels) were defined by Desikan-Killiany atlas24.
All processing steps except for step 2, alongside associated visualization, were fulfilled using the MRtrix3 package (www.mrtrix.org). The two connectomes were also imported into MATLAB for further analysis and comparison.
Results
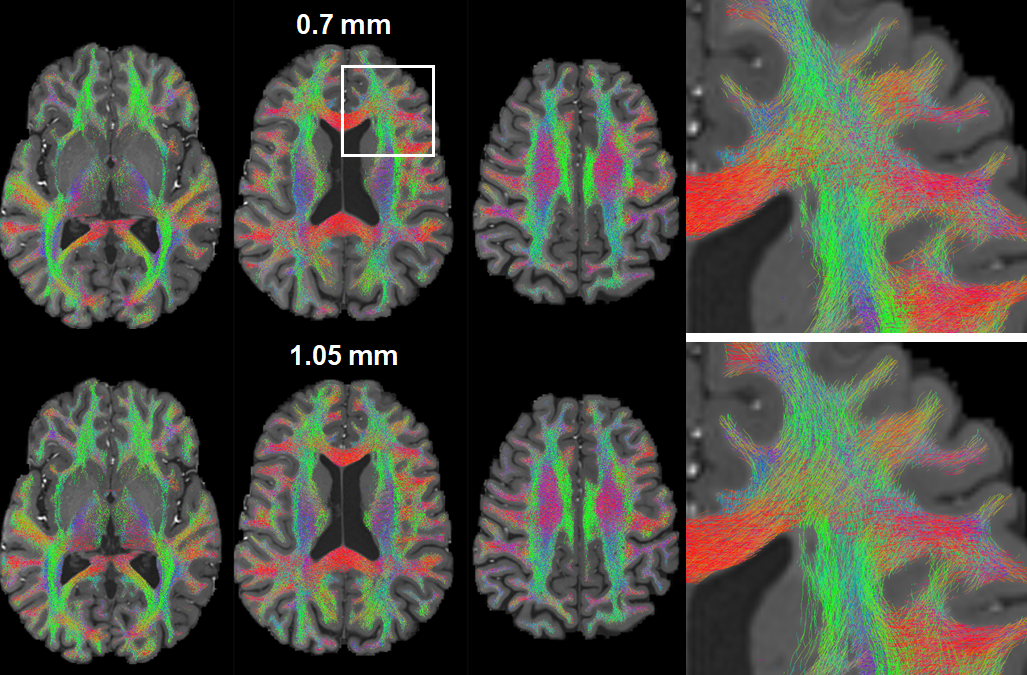
Despite the highly SNR-inefficient TR, the fODFs map estimated from the 0.7-mm diffusion images shared similar characteristic with that from the reference 1.05-mm data, effectively unraveling the fiber orientations in known anatomy (Fig. 2).
Correspondingly, the two resulting whole-brain tractograms presented a global pattern similar to each other (Fig. 3), both successfully delineating crossing fiber bundles as clearly identified when comparing exemplar zoom-ins.
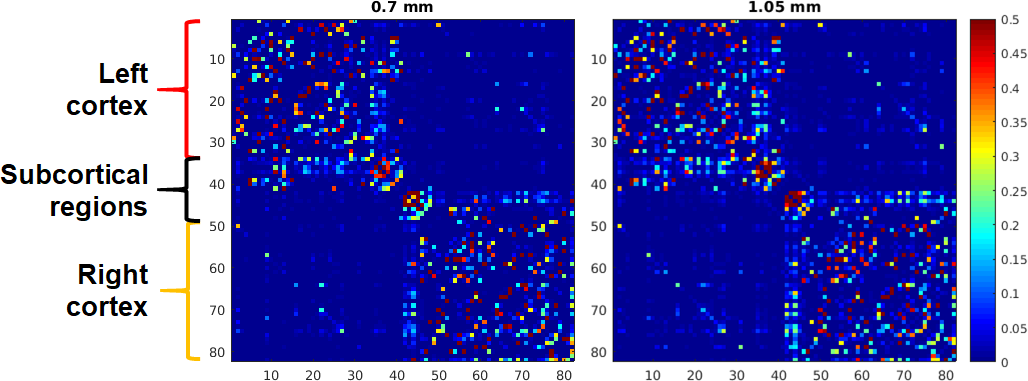
The similarity in tractograms in turn translated into two similar connectomes (Fig. 4) sharing the same characteristics such as strong intra-hemispheric cortical connections (correlation coefficient (cc): 0.96 for the left and 0.93 for the right hemisphere), noticeable homologue connections (cc: 0.87), and strong connections with subcortical regions (cc: 0.77).
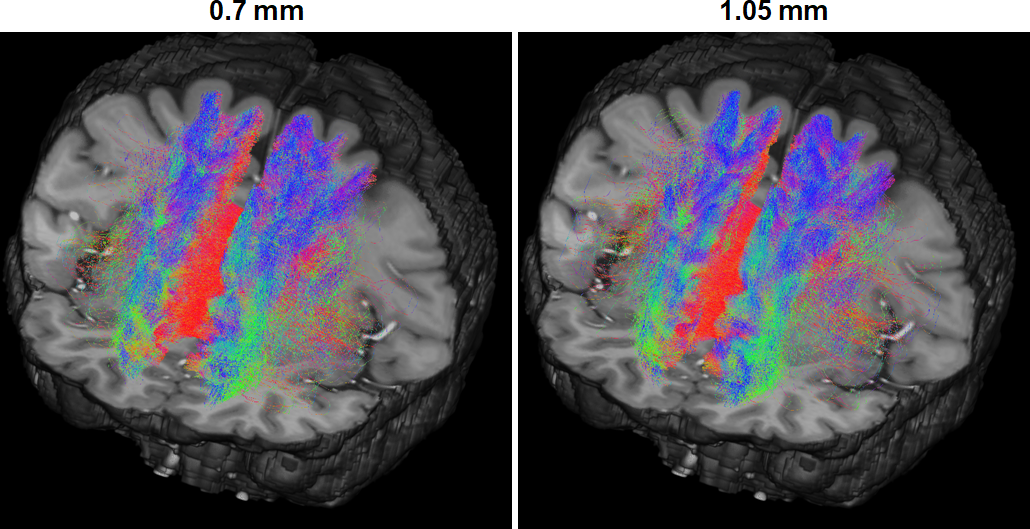
The similarity in connectome was further confirmed by inspecting selected streamlines connecting exemplar atlas regions: the left and right superior frontal regions (Fig. 5); the streamlines derived from 0.7-mm data characterized the homologue connection as well as that of the reference 1.05-mm data, effectively outlining how the two homologue regions are connected via corpus callosum.
Discussion and Conclusion
We have demonstrated that the SMS imaging can be used to accomplish submillimeter whole-brain diffusion at 7T with a standard body gradient.
Critical to this accomplishment was a synergistic combination of various techniques including pTx, FLEET ACS and denoising. Significant improvements are expected with a 64Rx coil enabling higher acceleration25.
Although largely improving the SNR, the current denoising strategy was found to not remove the noise effectively especially in subcortical regions (e.g., the thalamus).
Future work will examine alternative denoising strategies by concatenating multiple runs and/or removing the noise in the complex domain, the use of a 64Rx coil for improved SNR efficiency, and the utility of 0.7-mm diffusion for minimizing gyral bias26 and promoting the layer-specific analysis of intra-cortical architecture27,28.
Acknowledgements
The authors acknowledge Brian Hanna and John Strupp for setting up necessary computational resources. This work was supported by NIH grants U01 EB025144, P41 EB015894 and P30 NS076408.
References
1. Miller KL, Stagg CJ, Douaud G, Jbabdi S, Smith SM, Behrens TEJ, Jenkinson M, Chance SA, Esiri MM, Voets NL, Jenkinson N, Aziz TZ, Turner MR, Johansen-Berg H, McNab JA. Diffusion imaging of whole, post-mortem human brains on a clinical MRI scanner. NeuroImage 2011;57(1):167-181.
2. Chang HC, Sundman M, Petit L, Guhaniyogi S, Chu ML, Petty C, Song AW, Chen Nk. Human brain diffusion tensor imaging at submillimeter isotropic resolution on a 3Tesla clinical MRI scanner. NeuroImage 2015;118:667-675.
3. Setsompop K, Fan Q, Stockmann J, Bilgic B, Huang S, Cauley SF, Nummenmaa A, Wang F, Rathi Y, Witzel T, Wald LL. High-resolution in vivo diffusion imaging of the human brain with generalized slice dithered enhanced resolution: Simultaneous multislice (gSlider-SMS). Magnetic Resonance in Medicine 2018;79(1):141-151.
4. Moeller S, Yacoub E, Olman CA, Auerbach E, Strupp J, Harel N, Uǧurbil K. Multiband multislice GE-EPI at 7 tesla, with 16-fold acceleration using partial parallel imaging with application to high spatial and temporal whole-brain FMRI. Magnetic Resonance in Medicine 2010;63(5):1144-1153.
5. Setsompop K, Gagoski BA, Polimeni JR, Witzel T, Wedeen VJ, Wald LL. Blipped-controlled aliasing in parallel imaging for simultaneous multislice echo planar imaging with reduced g-factor penalty. Magnetic Resonance in Medicine 2012;67(5):1210-1224.
6. Uǧurbil K, Xu J, Auerbach EJ, Moeller S, Vu AT, Duarte-Carvajalino JM, Lenglet C, Wu X, Schmitter S, Van de Moortele PF, Strupp J, Sapiro G, De Martino F, Wang D, Harel N, Garwood M, Chen L, Feinberg DA, Smith SM, Miller KL, Sotiropoulos SN, Jbabdi S, Andersson JLR, Behrens TEJ, Glasser MF, Van Essen DC, Yacoub E. Pushing spatial and temporal resolution for functional and diffusion MRI in the Human Connectome Project. NeuroImage 2013;80:80-104.
7. Harms MP, Somerville LH, Ances BM, Andersson J, Barch DM, Bastiani M, Bookheimer SY, Brown TB, Buckner RL, Burgess GC, Coalson TS, Chappell MA, Dapretto M, Douaud G, Fischl B, Glasser MF, Greve DN, Hodge C, Jamison KW, Jbabdi S, Kandala S, Li X, Mair RW, Mangia S, Marcus D, Mascali D, Moeller S, Nichols TE, Robinson EC, Salat DH, Smith SM, Sotiropoulos SN, Terpstra M, Thomas KM, Tisdall MD, Ugurbil K, van der Kouwe A, Woods RP, Zollei L, Van Essen DC, Yacoub E. Extending the Human Connectome Project across ages: Imaging protocols for the Lifespan Development and Aging projects. Neuroimage 2018;183:972-984.
8. Wu X, Auerbach EJ, Vu AT, Moeller S, Lenglet C, Schmitter S, Van de Moortele P-F, Yacoub E, UÄŸurbil K. High-resolution whole-brain diffusion MRI at 7T using radiofrequency parallel transmission. Magnetic Resonance in Medicine 2018;80(5):1857-1870.
9. Vu AT, Auerbach E, Lenglet C, Moeller S, Sotiropoulos SN, Jbabdi S, Andersson J, Yacoub E, Ugurbil K. High resolution whole brain diffusion imaging at 7T for the Human Connectome Project. NeuroImage 2015;122:318-331.
10. Caruyer E, Lenglet C, Sapiro G, Deriche R. Design of multishell sampling schemes with uniform coverage in diffusion MRI. Magnetic Resonance in Medicine 2013;69(6):1534-1540.
11. Polimeni JR, Bhat H, Witzel T, Benner T, Feiweier T, Inati SJ, Renvall V, Heberlein K, Wald LL. Reducing sensitivity losses due to respiration and motion in accelerated echo planar imaging by reordering the autocalibration data acquisition. Magnetic Resonance in Medicine 2016;75(2):665-679.
12. Cauley SF, Polimeni JR, Bhat H, Wald LL, Setsompop K. Interslice leakage artifact reduction technique for simultaneous multislice acquisitions. Magnetic Resonance in Medicine 2014;72(1):93-102.
13. Sotiropoulos SN, Moeller S, Jbabdi S, Xu J, Andersson JL, Auerbach EJ, Yacoub E, Feinberg D, Setsompop K, Wald LL, Behrens TEJ, Ugurbil K, Lenglet C. Effects of image reconstruction on fiber orientation mapping from multichannel diffusion MRI: Reducing the noise floor using SENSE. Magnetic Resonance in Medicine 2013;70(6):1682-1689.
14. Veraart J, Novikov DS, Christiaens D, Ades-aron B, Sijbers J, Fieremans E. Denoising of diffusion MRI using random matrix theory. NeuroImage 2016;142:394-406.
15. Glasser MF, Sotiropoulos SN, Wilson JA, Coalson TS, Fischl B, Andersson JL, Xu J, Jbabdi S, Webster M, Polimeni JR, Van Essen DC, Jenkinson M. The minimal preprocessing pipelines for the Human Connectome Project. NeuroImage 2013;80(0):105-124.
16. Jeurissen B, Tournier JD, Dhollander T, Connelly A, Sijbers J. Multi-tissue constrained spherical deconvolution for improved analysis of multi-shell diffusion MRI data. NeuroImage 2014;103:411-426.
17. Tournier JD, Calamante F, Gadian DG, Connelly A. Direct estimation of the fiber orientation density function from diffusion-weighted MRI data using spherical deconvolution. NeuroImage 2004;23(3):1176-1185.
18. Dhollander T, Raffelt D, Connelly A. Accuracy of response function estimation algorithms for 3-tissue spherical deconvolution of diverse quality diffusion MRI data. Proc Intl Soc Mag Reson Med. Volume 26. Paris, France2018. p 1569.
19. Tournier J-D, Calamante F, Connelly A. Improved probabilistic streamlines tractography by 2nd order integration over fibre orientation distributions. Proc Intl Soc Mag Reson Med. Volume 18. Stockholm, Sweden2010. p 1670.
20. Smith RE, Tournier J-D, Calamante F, Connelly A. Anatomically-constrained tractography: Improved diffusion MRI streamlines tractography through effective use of anatomical information. NeuroImage 2012;62(3):1924-1938.
21. Smith RE, Tournier J-D, Calamante F, Connelly A. SIFT2: Enabling dense quantitative assessment of brain white matter connectivity using streamlines tractography. NeuroImage 2015;119:338-351.
22. Smith RE, Tournier J-D, Calamante F, Connelly A. The effects of SIFT on the reproducibility and biological accuracy of the structural connectome. NeuroImage 2015;104:253-265.
23. Hagmann P, Cammoun L, Gigandet X, Meuli R, Honey CJ, Van Wedeen J, Sporns O. Mapping the structural core of human cerebral cortex. PLoS Biology 2008;6(7):1479-1493.
24. Desikan RS, Ségonne F, Fischl B, Quinn BT, Dickerson BC, Blacker D, Buckner RL, Dale AM, Maguire RP, Hyman BT, Albert MS, Killiany RJ. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. NeuroImage 2006;31(3):968-980.
25. Moeller S, Grant A, Wu X, DelaBarre L, Van de Moortele P-F, Radder J, Schillak S, Auerbach EJ, Adriany G, Ugurbil K. Evaluation of Parallel Imaging performance gains with 64 channel receivers at 7 Tesla. 2018; Paris. p 141-141.
26. Sotiropoulos SN, Hernández-Fernández M, Vu AT, Andersson JL, Moeller S, Yacoub E, Lenglet C, Ugurbil K, Behrens TEJ, Jbabdi S. Fusion in diffusion MRI for improved fibre orientation estimation: An application to the 3T and 7T data of the Human Connectome Project. NeuroImage 2016;134:396-409.
27. Leuze CWU, Anwander A, Bazin PL, Dhital B, Stüber C, Reimann K, Geyer S, Turner R. Layer-specific intracortical connectivity revealed with diffusion MRI. Cerebral Cortex 2014;24(2):328-339.
28. Gulban OF, De Martino F, Vu AT, Yacoub E, Ugurbil K, Lenglet C. Cortical fibers orientation mapping using in-vivo whole brain 7 T diffusion MRI. Neuroimage 2018;178:104-118.
Figures