1042
White matter microstructure changes as a function of time-of-day: a biomarker of accumulating sleep pressure?1Department of Diagnostic Physics, Oslo University Hospital, Oslo, Norway, 2Norwegian Centre for Mental Disorders Research, Oslo University Hospital, Oslo, Norway
Synopsis
There is an increasing awareness of time-of-day (TOD) effects in MRI. The underlying causes of these fluctuations may be related to diurnal physiological variations, reflecting the processes that govern transition between sleep and wake. We probed TOD effects in a prospective manner, scanning 47 healthy individuals in the morning and the same evening using a five-shell diffusion-weighted imaging protocol at 3T, where a number of important Zeitgeber signals were rigorously controlled. We found significant changes in an number of derived DWI parameters (FA, MD, AWF, MK), and speculate that these changes are pointing towards underlying mechanisms of sleep-wake homeostasis.
Introduction
The role of diurnal fluctuations in the human brain derived by magnetic resonance imaging (MRI) is receiving increasing attention. However, little is known about the physiological changes underlying these fluctuations. Transition between stages of sleep and wake is regulated by circadian and homeostatic processes1. Contemporary theories of sleep regulation emphasise accumulation of learning-induced increases in synaptic strength2 and metabolic waste3 during wakefulness, which sleep then serves to renormalise. Diurnal fluctuations in MRI measures may thus reflect sleep regulating processes, and as such represent MRI-derived biomarkers of sleep pressure. Recent evidence suggests that structural measures by diffusion tensor imaging (DTI)4 change as a function of time-of-day (TOD), both in the white matter (WM) skeleton5, the whole cerebrum6 and at the interface of grey matter and cerebrospinal fluid (CSF)7. However, these studies lack rigorous experimental control between scans and only Elvsåshagen and colleagues5 assessed changes spanning more than a few hours. In contrast, the current study addressed the impact of TOD from morning to evening in a prospective controlled within-subject experimental design in which participants were continuously monitored from the first to the second scan. Moreover, the current study included estimations of non-Gaussian diffusion metrics obtained from diffusion kurtosis imaging (DKI) 8 and WM tract integrity9 in addition to classic DTI-derived parameters, in order to probe underlying axonal properties.Methods
Participants were 47 healthy adults (30 women) who underwent diffusion-weighted (DWI) MRI in the morning and evening of the same day. The imaging protocol consisted of a multi-shell full-brain multiband-accelerated 2 mm isotropic acquisition with b [0, 500, 1000, 2000, 3000]; directions [5, 12, 30, 40, 50]. Between scans, they stayed at the hospital premises under constant supervision and followed a standardised protocol of physical activity, food and fluid intake. Diffusion data were preprocessed in accordance with the following steps: noise correction10, Gibbs ringing corrections11, topup/eddy application12, B1 field normalisation and spatial smoothing with Gaussian kernet 1mm3. All diffusion metrics were then estimated using the Veraart and colleagues approach13. MRI images were analysed with voxel-wise tract based spatial statistics14, threshold free cluster enhancement15 and permutation testing in FSL16,17, correcting for multiple comparisons and thresholded at p < .05.Results
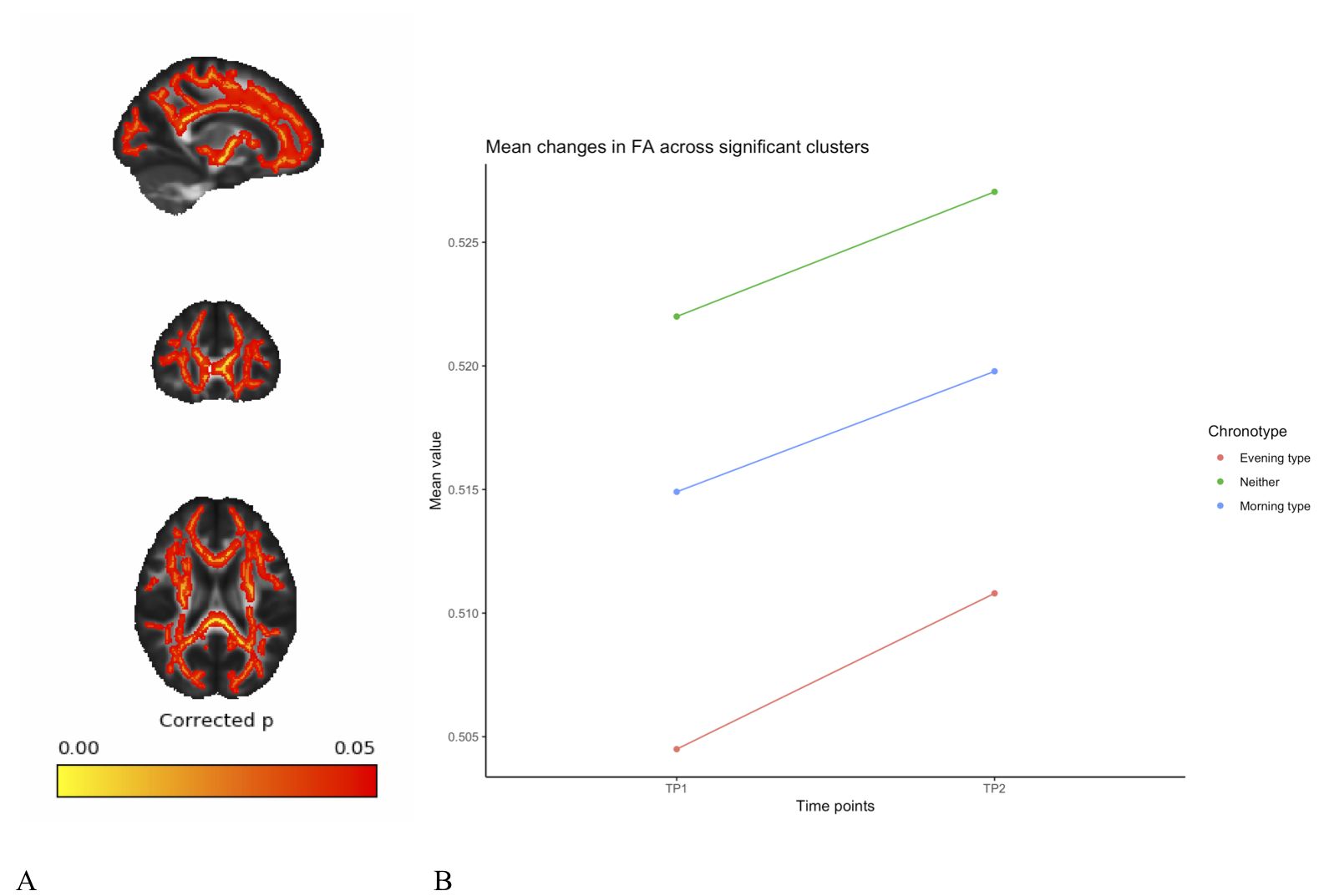
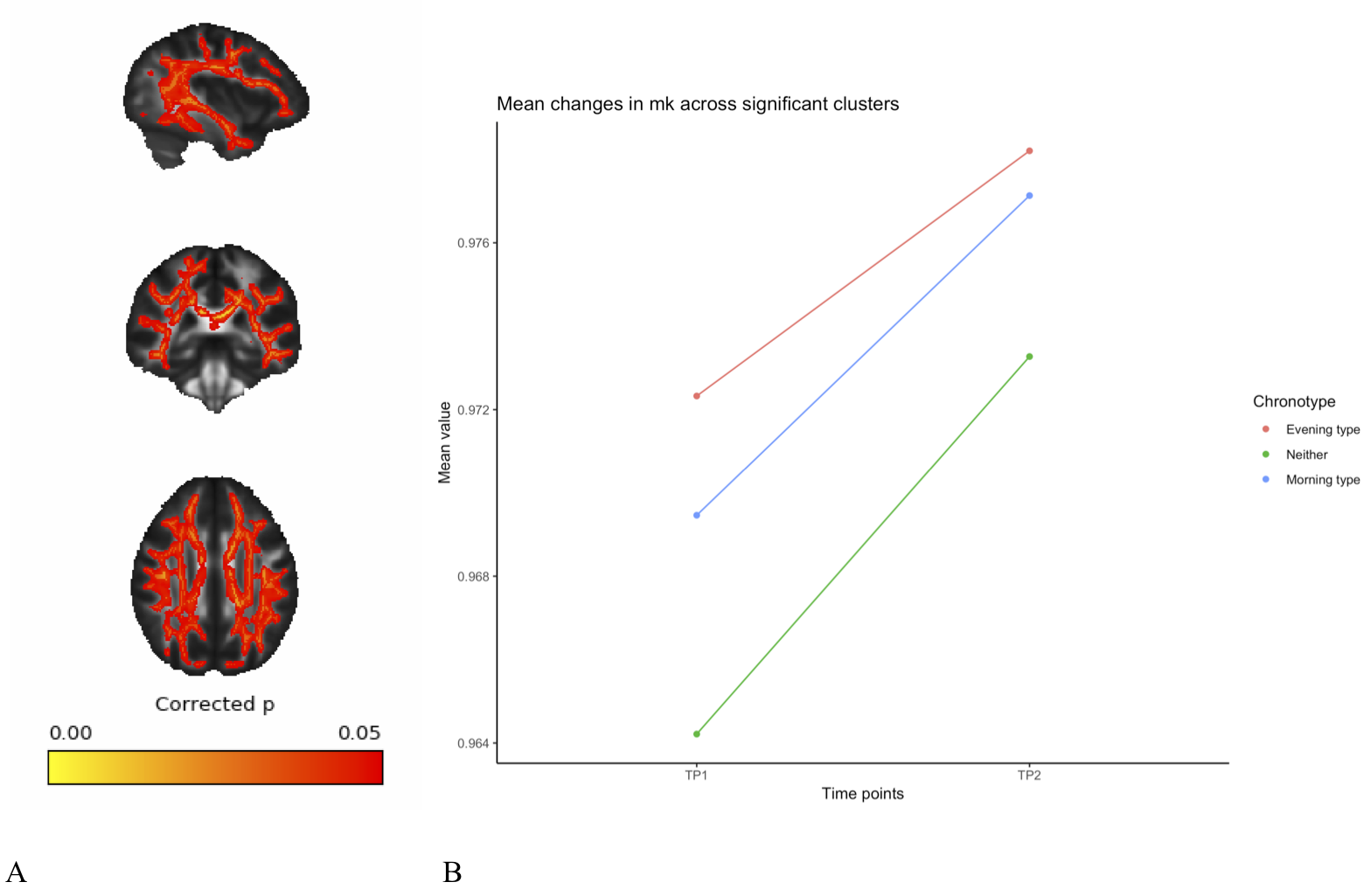
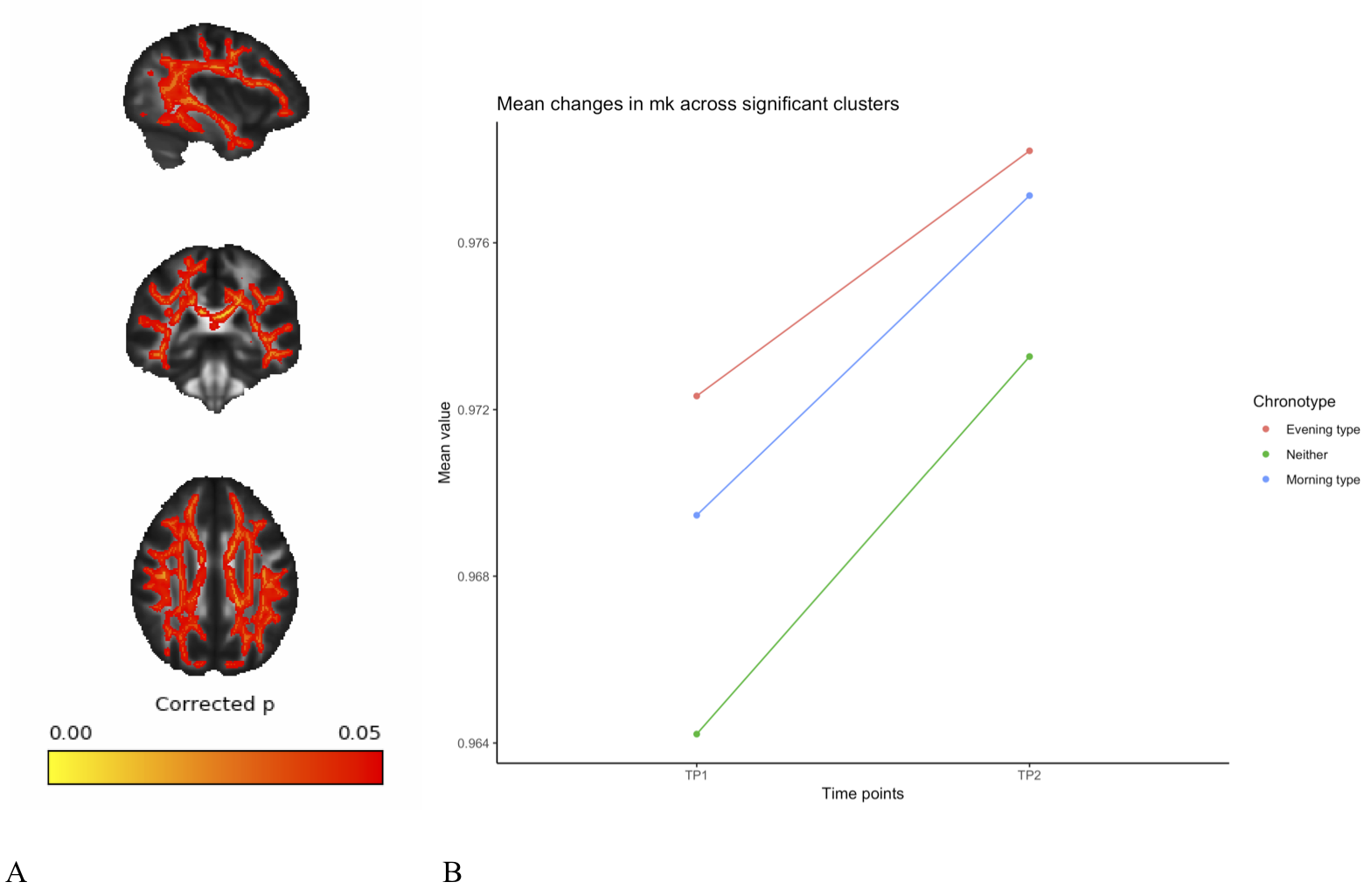
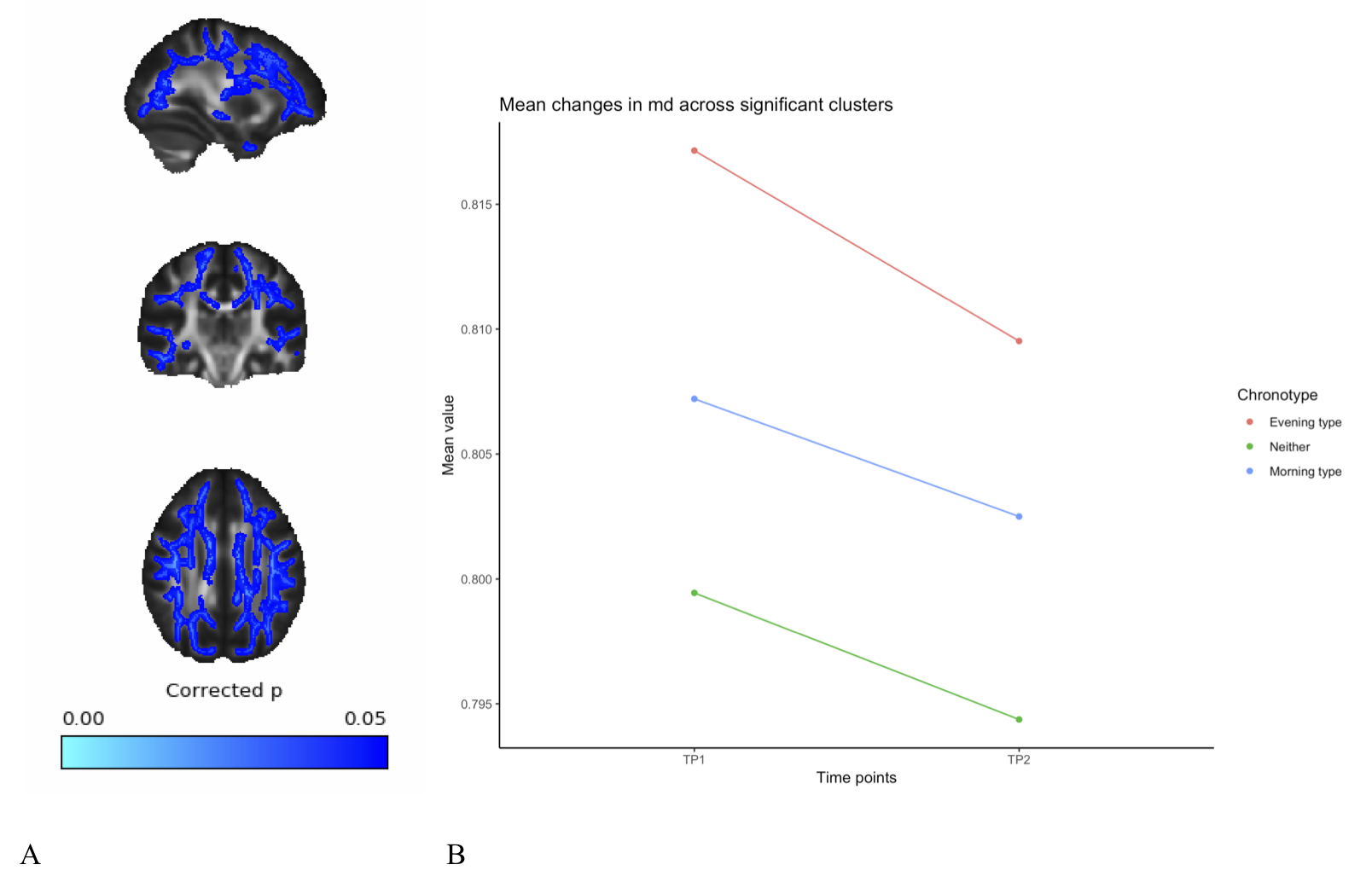
A full day of wakefulness was found to be associated with widespread increases in fractional anisotropy (FA), mean kurtosis (MK) and axonal water fraction (AWF), in addition to widespread decreases in mean diffusivity (MD). See figures 1-4. These changes were adjusted for age, gender and chronotype. Chronotype did not show a significant relationship with changes in any of the diffusion parameters. Moreover, changes spanned most of the WM skeleton, suggesting a global effect.Discussion
These findings indicate a temporal component to DWI-derived structural measures, in which white matter microstructure fluctuate as a function of TOD. These fluctuations may reflect physiological changes underlying accumulating sleep pressure, as the fluctuation is prevalent as a global change in WM microstructure. Importantly, we found an increase in estimations of WM integrity and specifically in axonal density, which suggest wakefulness may accommodate activity-dependent structural plasticity in WM. Specifically, this could reflect processes related to the activity-dependent accumulation of synapse strength2 and metabolic waste3. However, caution must be exerted in interpreting specific underlying biology on the basis of DTI and DKI18. Although the increases in the estimation of axonal density from AWF is a promising preliminary finding, introducing more complex diffusion models will afford more detailed investigation of this relationship. Despite this, these findings highlight the potential of MRI derived measures to probe sleep regulating processes in the human brain. Moreover, these findings have important implications for experimental considerations, as diurnal fluctuations would confound any design in which one experimental group is consistently scanned at a different TOD than another experimental group.Conclusion
The WM microstructure of the human brain is not static and fluctuate as a function of TOD. The fluctuations detected by DWI parameters may represent biomarkers of accumulating sleep pressure.Acknowledgements
We would like to thank the great team who helped with data collection.References
1. Borbély AA, Daan S, Wirz-Justice A, Deboer T. The two-process model of sleep regulation: A reappraisal. J Sleep Res 2016;25(2):131–43.
2. Tononi G, Cirelli C. Sleep and synaptic homeostasis: A hypothesis. Brain Res Bull 2003;62(2):143–50.
3. Jessen NA, Finmann Munk AS, Lundgaard I, Nedergaard M. The Glymphatic System – A Beginner’s Guide. Neurochem Res 2015;40(12):2583–99.
4. Basser PJ, Mattiello J, Le Bihan D. MR diffusion tensor spectroscopy and imaging. Biophys J 1994;66(1):259–67.
5. Elvsåshagen T, Norbom LB, Pedersen P, et al. Widespread changes in white matter microstructure after a day of waking and sleep deprivation. PLoS One 2015;10(5):1–15.
6. Jiang C, Zhang L, Zou C, et al. Diurnal microstructural variations in healthy adult brain revealed by diffusion tensor imaging. PLoS One 2014;9(1):1–10.
7. Thomas C, Sadeghi N, Nayak A, et al. Impact of time-of-day on diffusivity measures of brain tissue derived from diffusion tensor imaging. Neuroimage [Internet] 2018 [cited 2018 Apr 26];173:25–34. Available from: https://www.sciencedirect.com/science/article/pii/S1053811918301149
8. Jensen JH, Helpern JA, Ramani A, Lu H, Kaczynski K. Diffusional kurtosis imaging: The quantification of non-gaussian water diffusion by means of magnetic resonance imaging. Magn Reson Med [Internet] 2005 [cited 2018 Oct 27];53(6):1432–40. Available from: http://doi.wiley.com/10.1002/mrm.20508
9. Fieremans E, Jensen JH, Helpern JA. White matter characterization with diffusional kurtosis imaging. Neuroimage [Internet] 2011 [cited 2018 Oct 27];58(1):177–88. Available from: http://www.ncbi.nlm.nih.gov/pubmed/21699989
10. Veraart J, Novikov DS, Christiaens D, Ades-aron B, Sijbers J, Fieremans E. Denoising of diffusion MRI using random matrix theory. Neuroimage [Internet] 2016 [cited 2018 Oct 30];142:394–406. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27523449
11. Kellner E, Dhital B, Kiselev VG, Reisert M. Gibbs-ringing artifact removal based on local subvoxel-shifts. Magn Reson Med [Internet] 2016 [cited 2018 Oct 30];76(5):1574–81. Available from: http://www.ncbi.nlm.nih.gov/pubmed/26745823
12. Andersson JLR, Sotiropoulos SN. An integrated approach to correction for off-resonance effects and subject movement in diffusion MR imaging. Neuroimage [Internet] 2016;125:1063–78. Available from: http://dx.doi.org/10.1016/j.neuroimage.2015.10.019
13. Veraart J, Sijbers J, Sunaert S, Leemans A, Jeurissen B. Weighted linear least squares estimation of diffusion MRI parameters: Strengths, limitations, and pitfalls. Neuroimage [Internet] 2013 [cited 2018 Oct 30];81:335–46. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23684865
14. Smith SM, Johansen-Berg H, Jenkinson M, et al. Acquisition and voxelwise analysis of multi-subject diffusion data with tract-based spatial statistics. Nat Protoc 2006;2(3):499–503.
15. Smith SM, Nichols TE. Threshold-free cluster enhancement: Addressing problems of smoothing, threshold dependence and localisation in cluster inference. Neuroimage [Internet] 2009;44(1):83–98. Available from: http://dx.doi.org/10.1016/j.neuroimage.2008.03.061
16. Smith SM, Jenkinson M, Woolrich MW, et al. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage 2004;23(SUPPL. 1):208–19.
17. Winkler AM, Ridgway GR, Webster MA, Smith SM, Nichols TE. Permutation inference for the general linear model. Neuroimage [Internet] 2014;92:381–97. Available from: http://dx.doi.org/10.1016/j.neuroimage.2014.01.060
18. Novikov DS, Kiselev VG, Jespersen SN. On modeling. Magn Reson Med 2018;79(6):3172–93.
Figures