1009
Effective medium theory of multiple diffusion encoding1Center for Functionally Integrative Neuroscience, Aarhus University, Aarhus, Denmark, 2Radiology, NYU School of Medicine, New York, NY, United States
Synopsis
We consider the effect of tissue heterogeneity on diffusion acquisitions with multiple encodings. Previously, these signals were analyzed in Gaussian compartments, or disconnected pores. Here we assume a more realistic situation where a compartment cannot be considered Gaussian (uniform) at finite diffusion times, and derive the 4th-order contributions in the diffusion weightings, that distinguish the double-diffusion-encoding (DDE) signal from its single-encoding counterpart. We specifically identify terms odd in the DDE diffusion wave-vectors, which emerge due to local tissue heterogeneity but absent when compartments are Gaussian. Our results are expressed via the dynamical exponent related to the disorder universality class, and agree with Monte Carlo simulations.
Purpose
How different is a multiple-diffusion-encoding (MDE) from a set of single-diffusion encodings (SDE)? This fundamental question underpins currently popular applications of MDE sequences1-7. Here we answer this question up to $$${\cal\,O}(q^4)$$$, by accounting for the signature MDE contributions within the effective medium theory framework8,9. Introduction and overview of the results
Introduction and overview of the results
The conventional SDE signal10
$$S_t(q)=\int\frac{d^d r_1 d^d r_0}V\,{\cal G}_{t}(r_1,r_0)\,e^{-iq(r_1-r_0)}\,\equiv\,G(t,q)\,,\quad\,G(t,r)=\frac1V\int d^dr_0{\cal G}_{t}(r_0+r,r_0)$$
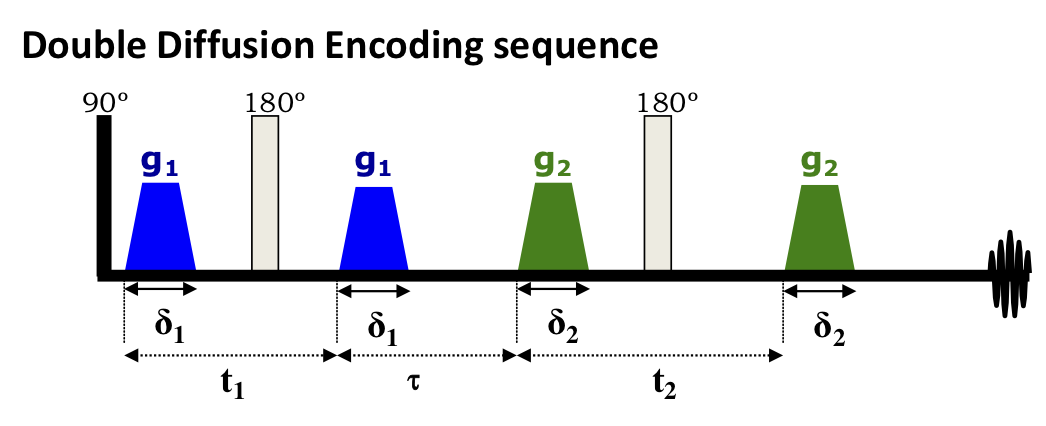
is given by the Fourier transform of the propagator $$$G=\langle{\cal G}\rangle$$$ integrated (i.e., averaged) over the medium due to summing over all diffusing spins in a voxel. For multiple-diffusion-encodings (MDE), such a simple relation does not hold11: Already for double-diffusion-encoding (DDE), Fig.1,12
$$S_{t_1,\tau,t_2}(q_1,q_2)=\int\! \frac{d{r_{1a}}d{r_{1b}}d{r_{2a}}d{r_{2b}}}V e^{ i{q_1}\cdot ({r_{1a}} - {r_{1b}})+i{q_2}\cdot ({r_{2a}}-{r_{2b}})}\,{{\cal G}_{{t_2};{r_{2b}},{r_{2a}}}} {{\cal G}_{\tau;{r_{2a}},{r_{1b}}}}{{\cal G}_{{t_1};{r_{1b}},{r_{1a}}}}\neq{G_{{t_2};{q_2}}} G_{\tau; 0} {G_{{t_1};{q_1}}}$$
the average of the product of exact local propagators $$$\langle{\cal G}{\cal G}{\cal G}\rangle$$$ does not equal the product of their medium-averages $$$GGG$$$ on the right-hand side. Therefore MDE gives new information starting from $$${\cal\,O}(q^4)$$$.13 So far, the deviations of MDE from SDEs was considered up to the 4th-order cumulant, $$$q^4$$$, for multiple Gaussian compartments, and disconnected pores in the long-time limit.11,13-15
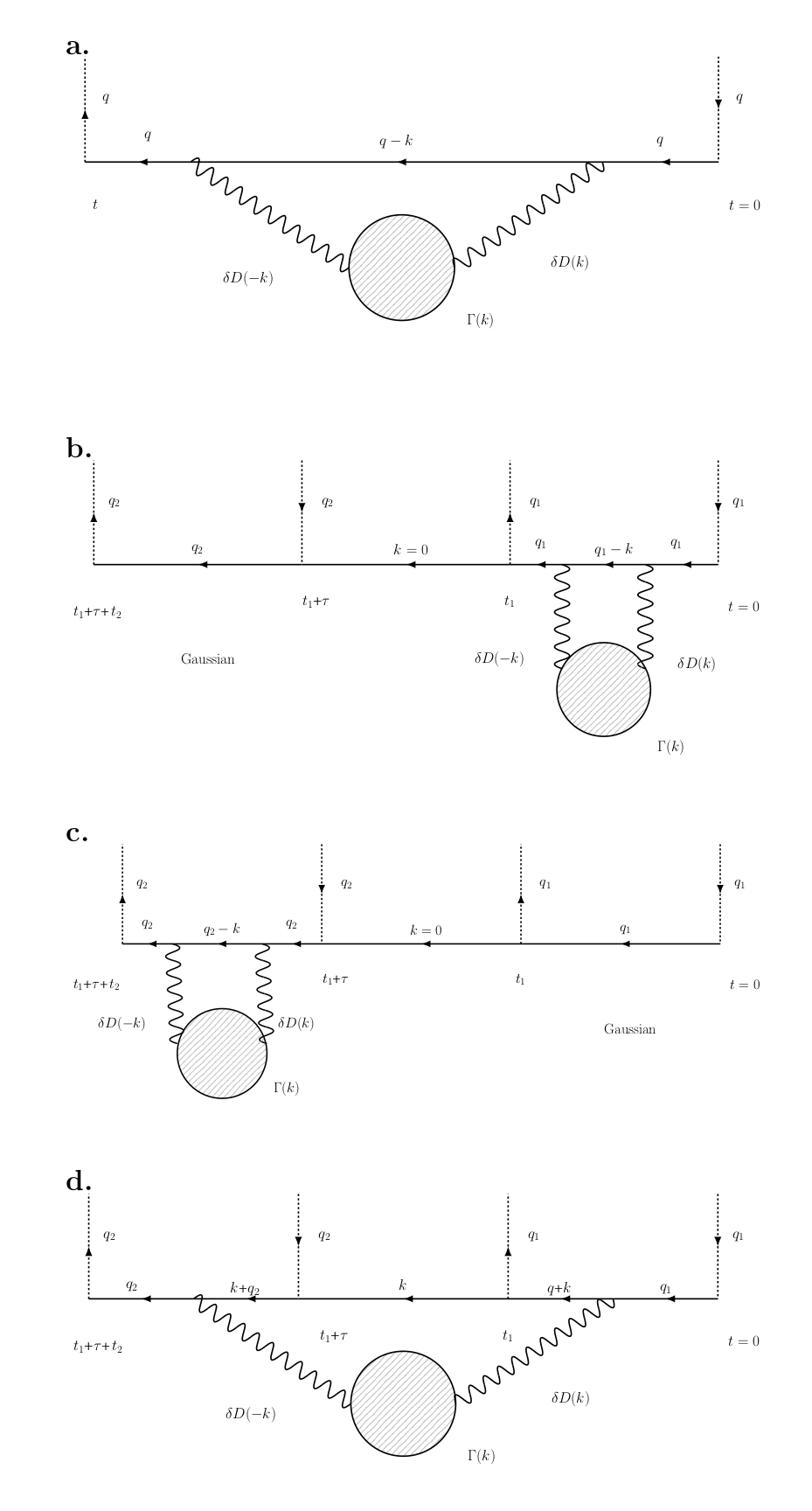
Here we consider a realistic tissue compartment where diffusion is non-Gaussian due to structural heterogeneity (e.g., hindrance in extra-cellular space; structural disorder in the intra-axonal space). We find the DDE signal, Fig.1, up to 4th order in $$$q_1$$$ and $$$q_2$$$. While the terms $$$q_1^4$$$ and $$$q_2^4$$$ appear as conventional kurtosis contributions on either of the DDE blocks, here we focus on the DDE-specific cross-terms $$$\sim\,q_1^2q_2^2$$$, as well as $$$\sim\,q_1q_2^3$$$, $$$q_1^3q_2$$$, in the cumulant expansion of $$$\ln\,S_{t_1,\tau,t_2}(q_1,q_2)$$$, depicted diagrammatically in Fig.2.
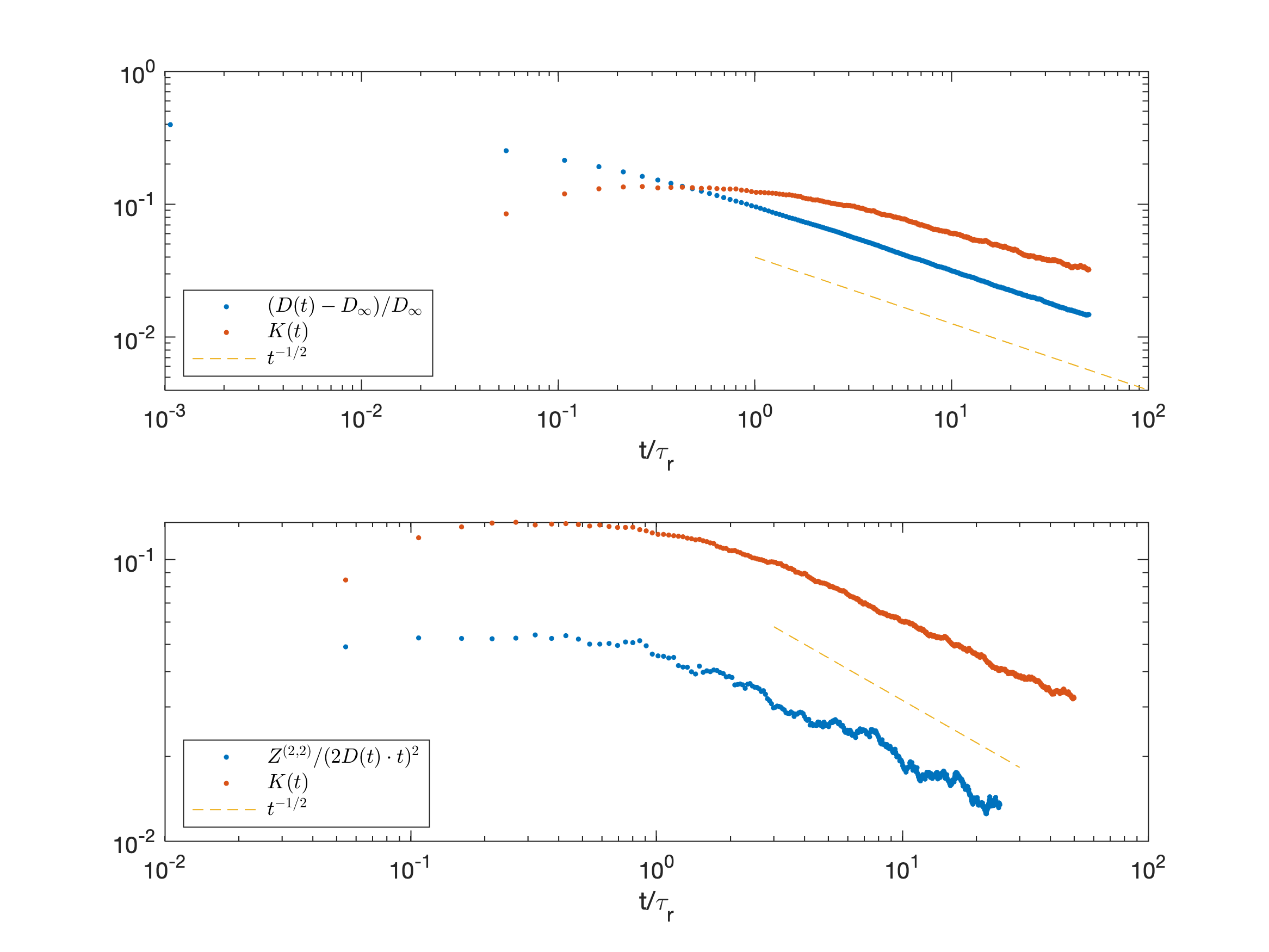
We show that these cross-terms reveal universal long-time scaling $$$\sim\,t^{2-\vartheta}$$$ characterized by the dynamical exponent $$\vartheta=\frac{p+d}2\qquad(1)$$ related to the structural exponent $$$p$$$ in $$$d$$$ dimensions9, and confirm it with $$$d=1$$$ simulations, Fig.3.
Cumulant representation of DDE
We define the DDE cumulants by the Taylor series
$$\ln S_{t_1,\tau,t_2}(q_1,q_2)=\sum_{n,m}\sum_{\substack{i_1\ldots i_n \\ j_1\ldots j_m}} \frac1{n!} \frac1{m!}(-i)^{n+m}q_{1i_1}\ldots q_{1i_n} q_{2j_1}\ldots q_{2j_m} \cdot Z_{i_1 \ldots i_n j_1 \ldots j_m}^{(n,m)} (t_1,t_2,\tau)$$
where only even $$$n$$$ and $$$m$$$ terms contribute in the absence of flow. The cumulants of molecular displacements $$$R_1=r_{t_1}-r_{0}$$$ and $$$R_2=r_{t_1+t_2+\tau}-r_{t_1+\tau}$$$
$$Z_{i_1 \ldots i_n j_1 \ldots j_m}^{(n,m)} (t_1,t_2,\tau)=\langle R_{1,i_1}\dots R_{1,i_n} R_{2,j_1}\dots R_{2,j_m}\rangle_c$$
such that the DTI terms
$$Z_{ij}^{(2,0)}=2t_1 D_{ij}(t_1)\quad\mbox{and}\quad Z_{ij}^{(0,2)} = 2t_2 D_{ij}(t_2);$$
the DKI terms16
$$Z_{ijkl}^{(4,0)}=(2\bar{D}t_1)^2 W_{ijkl}(t_1)\quad\mbox{and}\quad Z_{ijkl}^{(0,4)}=(2\bar{D}t_2)^2 W_{ijkl}(t_2).$$
The cross-term tensors mixing the DDE blocks $$$q_1$$$ and $$$q_2$$$
$$Z_{ijkl}^{(2,2)} = \langle R_{1i}R_{1j} R_{2k}R_{2l}\rangle_c;\quad\mbox{and}\quad Z_{ijkl}^{(1,3)}=\langle R_{1i}R_{2j} R_{2k}R_{2l}\rangle_c,\quad Z_{ijkl}^{(3,1)}=\langle R_{1i}R_{1j} R_{1k}R_{2l}\rangle_c\qquad(2)$$
are the lowest-order tensors unique to DDE.13,15
Effective medium theory for the 4th-order cumulant
Effective medium theory for the 4th-order cumulant
We represent the tissue by its spatially varying coarse-grained12 diffusivity $$$D(r)=D_0+\delta D(r)$$$, asymptotically exact for long diffusion times.9 The structural disorder is characterized by the power spectrum
$$\Gamma(k)=\frac1V\langle\delta D(-k)\delta D(k)\rangle\sim k^p\,,\quad\,k\to0$$
where the structural exponent $$$p$$$ entering Eq.(1) determines the structural universality class of the disorder. The desired cross-terms, Eq.(2), can be found from the Feynman diagram in Fig.2d. While its exact expression is cumbersome, we give a few transparent limits.
Short-time limit: $$$t_1\to0,\,\,\,t_2\to0$$$
For zero mixing time $$$\tau=0$$$,
$$Z^{(2,2)}_{ijkl}\simeq 4t_1 t_2 \delta_{ij}\delta_{kl}\langle(\delta D)^2\rangle\,,\quad\,t_1,t_2\to0,$$
diffusing spins act as if residing in a multi-compartmental system characterized by the distribution of local diffusivities $$$D(r)$$$. Thus, it is only in this limit that the $$$b^2$$$ term of the DDE reflects the diffusion tensor covariance $$$\langle(\delta D)^2\rangle$$$.
For finite mixing time, this is generalized to
$$Z^{(2,2)}_{ijkl}\simeq4t_1 t_2\delta_{ij}\delta_{kl}\int \frac{d^d k}{(2\pi)^d} e^{-D_0k^2\tau}\;\Gamma(k),\quad\,t_1,t_2\to0,$$
i.e. Gaussian smoothing of the local diffusivity due to diffusion during the mixing time.
The "odd" $$$q_1q_2^3$$$ and $$$q_1^3q_2$$$ cross-terms are even more interesting:
$$Z_{ijkl}^{(1,3)}\simeq6t_1 t_2(D_0t_2)\delta_{ij}\delta_{kl}\int \frac{d^d k}{(2\pi)^d}e^{-D_0 k^2\tau}k^2\Gamma(k)\,,\quad\,t_1,t_2\to0,$$
again a Gaussian-filtered version of the $$$\tau=0$$$ result
$$Z_{ijkl}^{(1,3)}\simeq6t_1 t_2(D_0t_2)\delta_{ij}\delta_{kl}\langle(\nabla D)^2\rangle\,,\quad\,t_1,t_2\to0.$$
Interestingly, the short time behavior of $$$Z^{(1,3)}$$$ now reflects the correlation of the local diffusivity gradient, consistent with $$$Z_{ijkl}^{(1,3)}\equiv0$$$ in systems of multiple Gaussian compartments. A nonvanishing $$$Z^{(1,3)}$$$ tensor may be identified by its characteristic $$$\cos^3\psi$$$ behavior of the powder averaged DDE signal, as the angle $$$\psi$$$ between the wavevectors $$$q_1$$$ and $$$q_2$$$ is varied.
Long-time limit:$$$\quad\,t_1=t_2=t\to\infty;\quad\tau\to0$$$
We find universal scaling
$$Z^{(2,2)}_{ijkl}\sim t^{2-\vartheta}\,,\quad Z^{(1,3)}_{ijkl}\sim t^{2-\vartheta}\,,\quad Z^{(3,1)}_{ijkl}\sim t^{2-\vartheta}\qquad(3)$$
with known coefficients that change sign as function of $$$p$$$ and $$$d$$$. Such scaling behavior is similar to the kurtosis term $$$W\sim t^{-\vartheta}$$$, such that $$$Z^{(4,0)}\sim Z^{(0,4)}\sim t^{2-\vartheta}$$$, and to $$$-\ln S\sim t^{2-\vartheta}$$$ divergent ADC correction due to mesoscopic magnetic structure.17
Monte Carlo simulations in Fig.3 confirm the $$$t^{2-\vartheta}$$$ scaling in $$$d=1$$$ dimensional array of randomly placed permeable barriers.9
Conclusions
We developed effective medium theory for the time dependence of rank-4 tensors unique to DDE. We found that at short times, these tensors reflect the variance in local diffusivity and its gradient, whereas their $$$t\to\infty$$$ scaling is similar to the SDE cumulants. The characteristic behavior of $$$Z^{(1,3)}$$$ could make it a probe for detecting violations of the commonplace assumption of multiple Gaussian compartments. Our results point out limitations to current interpretations of MDE tensors.Acknowledgements
The authors are grateful to Antonius Papaioannou and Vinu Abeywick for technical assistance.
Research was supported by the Lundbeck foundation (R291-2017-4375), AUFF, and Augustinus foundation (18-1456), by R01 NS088040 grant from the NINDS (NIH), and by the Center of Advanced Imaging Innovation and Research (CAI2R, www.cai2r.net), a NIBIB Biomedical Technology Resource Center: P41 EB017183.
References
[1] Marco Lawrenz and Jurgen Finsterbusch. Detection of microscopic diffusion anisotropy in human cortical gray matter in vivo with double diffusion encoding. Magnetic resonance in medicine, Sep 2018.
[2] Andrada Ianus, Sune N. Jespersen, Teresa Serradas Duarte, Daniel C. Alexander, Ivana Drobnjak, and Noam Shemesh. Accurate estimation of microscopic diffusion anisotropy and its time dependence in the mouse brain. NeuroImage, 183:934–949, Dec 2018.
[3] Yang Ji, Jeffrey Paulsen, Iris Yuwen Zhou, Dongshuang Lu, Patrick Machado, Bensheng Qiu, Yi-Qiao Song, and Phillip Zhe Sun. In vivo microscopic diffusional kurtosis imaging with symmetrized double diffusion encoding EPI. Magnetic Resonance in Medicine, Sep 2018.
[4] Noam Shemesh, Jens T. Rosenberg, Jean-Nicolas Dumez, Samuel C. Grant, and Lucio Frydman. Distinguishing neuronal from astrocytic subcellular microstructures using in vivo double diffusion encoded 1H MRS at 21.1 T. PloS one, 12:e0185232, 2017.
[5] Grant Yang, Qiyuan Tian, Christoph Leuze, Max Wintermark, and Jennifer A Mcnab. Double Diffusion Encoding MRI for the Clinic. Magnenetic Resonance in Medicine, 520:507–520, Aug 2018.
[6] Filip Szczepankiewicz, Samo Lasic, Danielle van Westen, Pia C Sundgren, Elisabet Englund, Carl-Fredrik Westin, Freddy Stahlberg, Jimmy Latt, Daniel Topgaard, and Markus Nilsson. Quantification of microscopic diffusion anisotropy disentangles effects of orientation dispersion from microstructure: applications in healthy volunteers and in brain tumors. NeuroImage, 104:241–252, 2015.
[7] Jens H. Jensen, Edward S. Hui, and Joseph A. Helpern. Double-pulsed diffusional kurtosis imaging. NMR in Biomedicine, 27(4):363–370, 2014.
[8] Dmitry S. Novikov and Valerij G Kiselev. Effective medium theory of a diffusion-weighted signal. NMR Biomed, 23(7):682–697, Aug 2010.
[9] Dmitry S. Novikov, Jens H Jensen, Joseph A Helpern, and Els Fieremans. Revealing mesoscopic structural universality with diffusion. Proc Natl Acad Sci U S A, 111(14):5088–5093, Apr 2014.
[10] Paul T. Callaghan. Principles of nuclear magnetic resonance microscopy. Clarendon Press, 1993.
[11] Partha Mitra. Multiple wave-vector extensions of the NMR pulsed-field-gradient spin-echo diffusion measurement. Phys Rev B 51(21):15074–15078, Jun 1995.
[12] Dmitry S. Novikov, Sune N Jespersen, Valerij G Kiselev, and Els Fieremans. Quantifying brain microstructure with diffusion MRI: Theory and parameter estimation. NMR in Biomedicine 2018, DOI: 10.1002/nbm.3998
[13] Sune Nørhøj Jespersen. Equivalence of double and single wave vector diffusion contrast at low diffusion weighting. NMR in Biomedicine, 25(6):813–818, Jun 2012.
[14] Marco Lawrenz, Martin A. Koch, and Jurgen Finsterbusch. A tensor model and measures of microscopic anisotropy for double-wave-vector diffusion-weighting experiments with long mixing times. Journal of Magnetic Resonance 202:43–56, Jan 2010.
[15] Sune Nørhøj Jespersen, Henrik Lundell, Casper Kaae Sønderby, and Tim B. Dyrby. Orientationally invariant metrics of apparent compartment eccentricity from double pulsed field gradient diffusion experiments. NMR in Biomedicine, 26(12):1647–1662, 2013.
[16] Jens H Jensen, Joseph A Helpern, Anita Ramani, Hanzhang Lu, and Kyle Kaczynski. Diffusional kurtosis imaging: the quantification of non-gaussian water diffusion by means of magnetic resonance imaging. Magn Reson Med, 53(6):1432–1440, Jun 2005.
[17] Dmitry S. Novikov, Marco Reisert, and Valerij G. Kiselev. Effects of mesoscopic susceptibility and transverse relaxation on diffusion NMR. Journal of Magnetic Resonance, 293:134-144, 2018.
Figures