0911
Subject specific functional connectivity fingerprints made possible with temporal ICA1Donders Centre for Brain, Cognition and Behaviour, Radboud University, Nijmegen, Netherlands, 2MIRA Institute for Biomedical Technology and Technical Medicine, University of Twente, Enschede, Netherlands, 3Erwin L. Hahn Institute for Magnetic Resonance Imaging, Essen, Germany
Synopsis
In the current contribution we use an ultra-fast fMRI sequence (TR=0.158s) and temporal ICA to generate subject specific fingerprints of unprecedented specificity. These "fingerprints" are generated from the mappings between common RSNs and time-series of temporal ICs. We also show that these time-series are virtually free from physiological contamination.
Introduction
Resting-state fMRI has been used for the past twenty years as a tool to study functional connectivity (FC)1. Analysis techniques such as seed-based correlation analysis2 and spatial ICA3 have been used to identify common spatial patterns of activity now referred to as resting-state networks (RSNs). Nonetheless, because RSNs analysis suffer from issues ranging from dependency on pre-processing steps4, physiological noise and motion contamination5, and, possibly, "network splitting", the study of RSNs and FC has not been able to prove itself robust enough for neuroscientific nor clinical investigations (see, e.g., ref. 6).
In the current contribution we show that, by using an ultra-fast fMRI sequence7 and an analysis strategy based on temporal ICA and temporal functional modes8, the resulting decomposition can be used at the single-subject level to generate subject fingerprints of unprecedented specificity.
Methods
A multiband echo-shifted (MESH) EPI sequence was implemented to sample the BOLD signal with full brain coverage and homogenous resolution. The protocol was designed with the following parameters: TR = 158ms, TE = 48ms, resolution = 4.5mm, 30 slices without gap, SMS=6 with a CAIPI FOV/3 shift, no in-plane acceleration and no partial Fourier, no Fat Saturation and a flip angle of 8 degrees. We acquired 6 11-minute (4096 volumes) fMRI runs from 6 subjects, for a total of 36 datasets. Data were acquired on a 3T Siemens Prisma scanner using a 32-channel coil. Data were preprocessed using fMRIPREP and high-pass filtered (cutoff 1/100s). Each dataset was decomposed using FSL MELODIC into 120 spatial ICA components, which were subsequently classified into signal and noise. Preprocessed datasets were denoised by regressing out noise components with FSL regfilt.
To examine whether TFMs depend on the initial choice of template, three different temporal ICA strategies were implemented. The first strategy consisted of running a 21-dimensional temporal ICA directly on spatial ICA signal time-courses. For the second strategy, denoised datasets were used as inputs for a group 120-dimensional ICA decomposition, from which 59 signal components were identified. These 59 signal components served as a template for dual regression. Run-specific dual regression time-courses were finally unmixed using temporal ICA, yielding 21 independent time-courses. The third strategy consisted in extracting 444 time-series from each denoised dataset using an atlas based parcellation9, and then un-mixing those time-series.
To show that independent time-courses are not contaminated by confounds, we computed their correlation coefficient to 32 confound regressors obtained with FMRIPREP.
Finally, to examine the subject specific nature of the decomposition, we use a structure preserving dimensionality reduction technique10 to compare the topological structure of RSN and TFM time-series, as well as TFM core mixing matrices.
Results
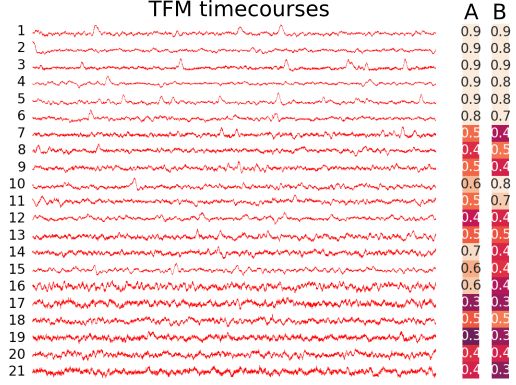
Figure 1 illustrates that independent timecourses are to a large extent independent on the original choice of data dimensionality reduction. According to the degrees of freedom of the initial parcellation, however, maps may look more or less smoothed, and there may be more or less mixtures to estimate the underlying latent signals.
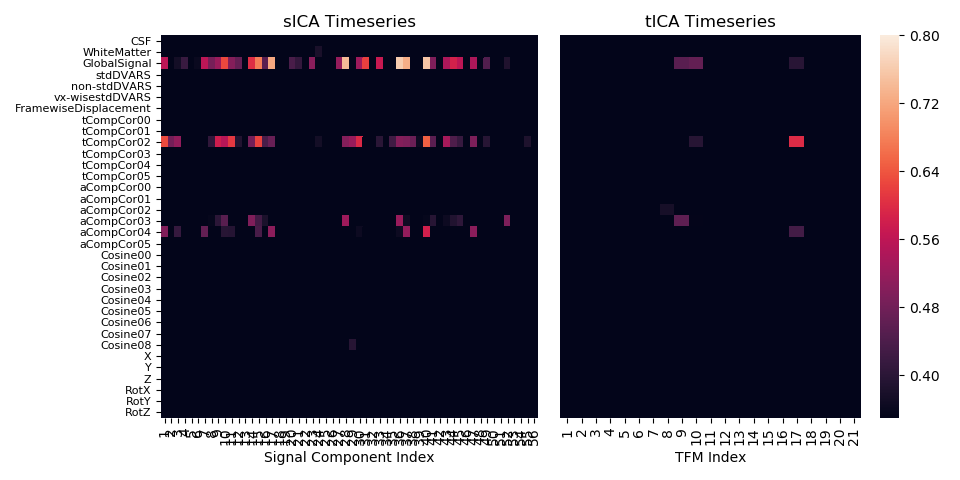
Figure 2 shows that independent timecourses are free from confounding contamination. A similar result focussing on the global signal, specifically, has been reported previously in the literature11.
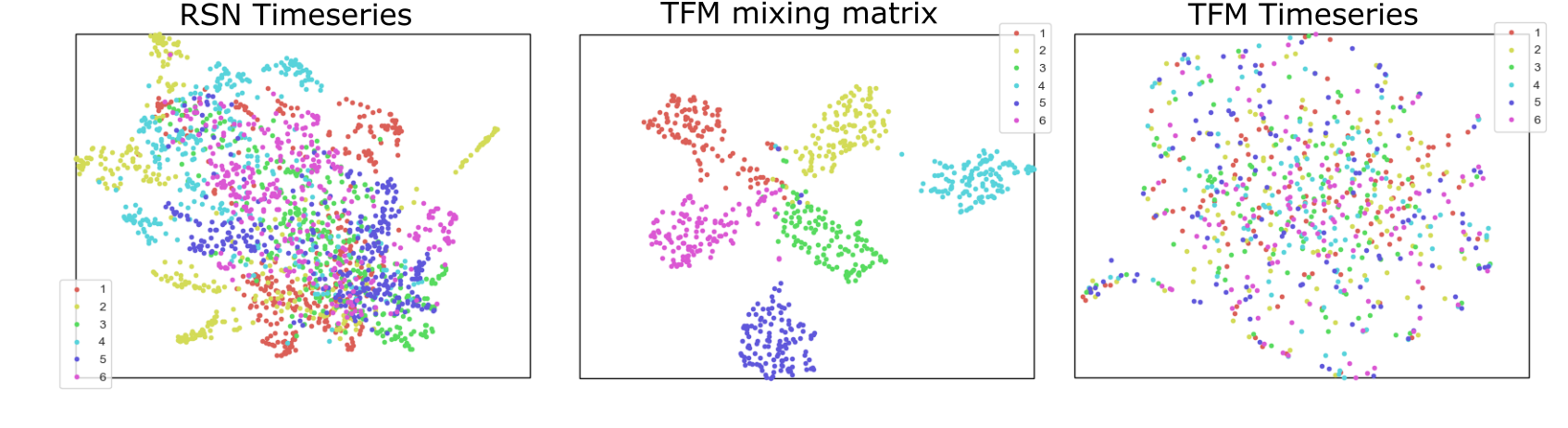
Figure 3 shows that the RSN to TFM mappings constitute subject fingerprints with unprecedented specificity. By unmixing RSN timeseries with temporal ICA it is possible to separate subject specific information (contained in the TFM mixing matrix) from the independent TFM timecourses (which are devoid of subject specificity).
Discussion
Because of the inherently slow sampling rates offered by traditional MRI sequences, and thus small number of time-points, temporal ICA nremains unexplored at the single subject level. Recent advances in MR technology, however, allow ultra-fast sampling of the BOLD signal with full brain coverage and isotropic resolution, such that thousands of time-points can be obtained within minutes. By taking advantage of an ultra-fast fMRI sequence we show that temporal ICA can be performed on the single-subject level, and it is capable of extracting exquisite subject specific information not yet seen in FC studies. We show here that problems of contamination associated with the study of RSN networks are not present in analyses based on temporal ICA. Additionally, we obtained highly separable subject specific fingerprints from RSN to TFM mappings, parsimoniously, without the need for feature engineering nor big data.
There is, however, much more left to be explored in the content of the TFM spatial maps (which indicate the interactions between RSNs), as well as in the independent time-courses, which are virtually free of contaminants and weakly dependent on initial data reduction strategies.
Acknowledgements
We thank Christian Beckmann for fruitful discussions. Daniel Gomez is funded by a Marie Curie FP7-PEOPLE-2013-ITN "Initial Training Networks" Action from the European Union (Project Reference Number: 608123).References
[1] M. P. van den Heuvel and H. E. Hulshoff Pol. “Exploring the brain network: a review on resting-state fMRI functional connectivity.” In: European neuropsychopharmacology : the journal of the European Col- lege of Neuropsychopharmacology 20.8 (2010), pp. 519–34.
[2] B. Biswal, F. Z. Yetkin, V. M. Haughton, and J. S. Hyde. “Functional connectivity in the motor cortex of resting human brain using echo- planar MRI”. In: Magn. Reson. Med. OCTOBER (1995).
[3] C. F. Beckmann, S. M. Smith, and F. M. Smith. “Probabilistic In- dependent Component Analysis for Functional Magnetic Resonance Imaging”. In: IEEE Transactions on Medical Imaging 23.1 (2004), pp. 137– 152.
[4] D. M. Cole, S. M. Smith, and C. F. Beckmann. “Advances and pit- falls in the analysis and interpretation of resting-state FMRI data”. In: Frontiers in Systems Neuroscience 4.April (2010), pp. 1–15.
[5] K. Murphy, R. M. R. Birn, and P. A. P. Bandettini. “Resting-state fMRI confounds and cleanup”. In: NeuroImage 80 (2013), pp. 349–359. arXiv: NIHMS150003.
[6] P. C. Mulders, P. F. van Eijndhoven, A. H. Schene, C. F. Beckmann, and I. Tendolkar. “Resting-state functional connectivity in major de- pressive disorder: A review”. In: Neuroscience and Biobehavioral Re- views 56 (2015), pp. 330–344.
[7] R. Boyacioğlu, J. Schulz, and D. G. Norris. “Multiband echo-shifted echo planar imaging”. In: Magnetic Resonance in Medicine 77.5 (2017), pp. 1981–1986. 6
[8] S. M. Smith, K. L. Miller, S. Moeller, J. Xu, E. J. Auerbach, M. W. Woolrich, C. F. Beckmann, M. Jenkinson, J. Andersson, M. F. Glasser, D. C. Van Essen, D. A. Feinberg, E. S. Yacoub, and K. Ugurbil. “Temporally-independent functional modes of spontaneous brain activity”. In: Proceedings of the National Academy of Sciences 109.8 (2012), pp. 3131–3136.
[9] S. Urchs, J. Armoza, Y. Benhajali, J. St-Aubin, P. Orban, and P. Bellec. “MIST: A multi-resolution parcellation of functional brain net- works”. In: MNI Open Research 1.0 (2017), p. 3.
[10] L. McInnes and J. Healy. “UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction”. In: (2018), pp. 1–18. arXiv: 1802.03426.
[11] M. F. Glasser, T. S. Coalson, J. Bijsterbosch, S. J. Harisson, M. P. Harms, A. Anticevic, D. C. Van Essen, and Stephen M. Smith. “Using Temporal ICA to Selectively Remove Global Noise While Preserving Global Signal in Functional MRI Data”. In: bioRxiv (2017).
Figures