0906
Capturing brain spatial topography reconfiguration using ultra-high-order independent component analysis (ICA)1The Mind Research Network, Albuquerque, NM, United States, 2School of Medicine, Yale University, New Haven, CT, United States, 3Department of Electrical and Computer Engineering, University of New Mexico, Albuquerque, NM, United States
Synopsis
The brain reorganizes its activity interactively at different temporal and spatial scales. These reconfigurations include variations within a region’s
Acronym
fMRI: Functional magnetic resonance imaging
ICA: independent component analysis
FU: functional unit
Introduction
There are ongoing neuroscience discussions on brain dynamics. Considering the prominent role of fMRI in our understanding of brain function and organization, there is a growing effort utilizing fMRI to capture brain dynamics. However, previous work has mainly focused on temporal variations among fixed spatial nodes, ignoring spatial reconfiguration over time. Recently, we developed a new approach to capture spatiotemporal variations of functional domains using the concept of functional hierarchy within the brain. The findings of this study demonstrate spatial reconfiguration of the brain’s networks and changes in a given brain region’s functional roles over time, highlighting the need to use data-driven approaches to detect functional units rather than current parcellations approaches. This abstract presents the latest progress of this ongoing project aiming to capture within and between-subject spatiotemporal variations via the estimation of brain functional units (FUs).Methods
Data collection was performed at 7 imaging sites across the United States. Details of the dataset and preprocessing procedure can be found in our earlier work1. First, low-order ICA (20 components) was applied to identify large-scale networks including the left and right frontoparietal (LPFN and RPFN), primary visual (VisPri), somatomotor (MTR), default mode (DM), cerebellar (CER), salience (SN), secondary visual (VisSec), auditory (AUD), attention (ATN), and language (LANG) networks. Ultra-high-order ICA (1000 components) was subsequently used to estimate FUs. An FU is a pattern of brain regions with very similar functional activity over time. FUs were assigned to networks using the similarity of temporal activity between FUs and brain networks. First, the dominant network assignment of each FU was obtained using the whole time series and all subjects (static group labeling). Next, for each time window (60 secs), FUs were assigned to networks using a novel regularized cost function. The cost function simultaneously maximizes the similarity between the temporal activities of a given FU and minimizes the divergence from the dominant assignment. Therefore, it estimates FU assignments to large-scale networks while protecting against the misassignment of FUs due to spurious fluctuations, and optimizing the selection of the best subsets of changes from group static assignment.Results
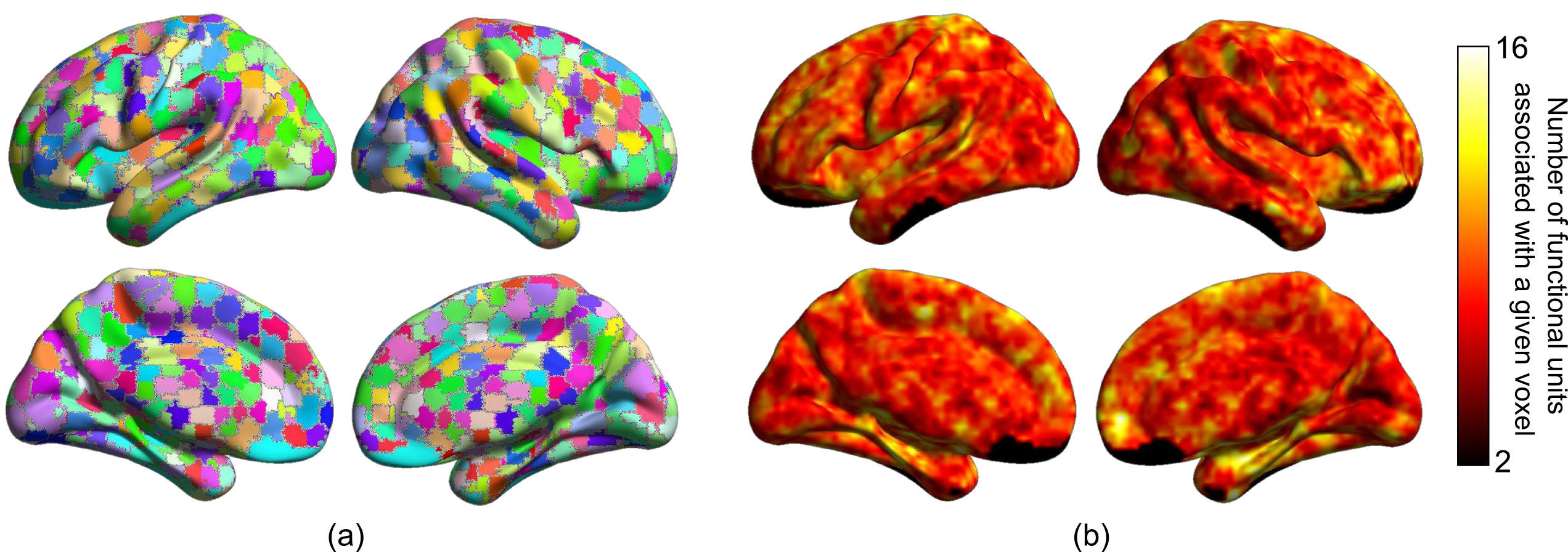
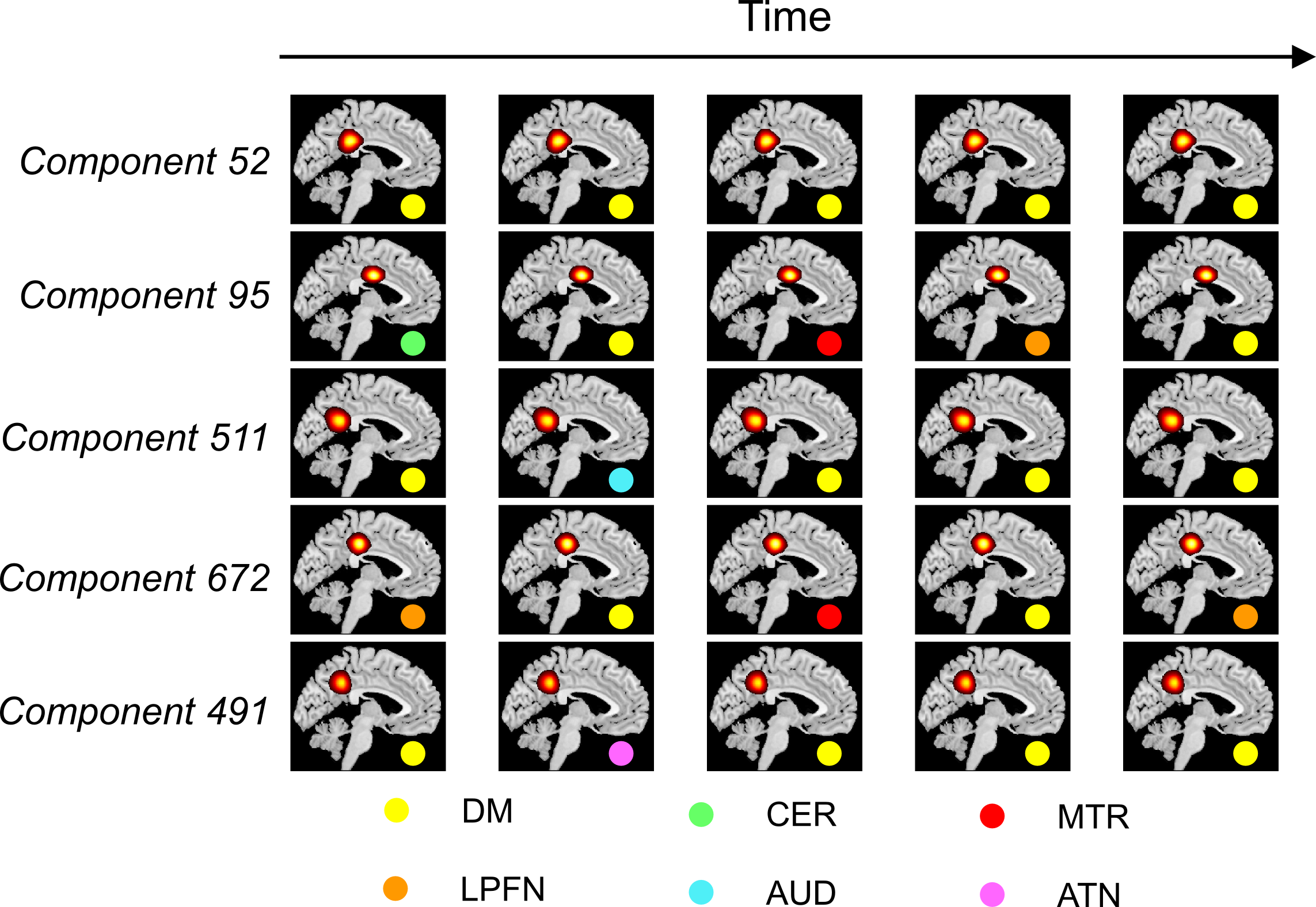
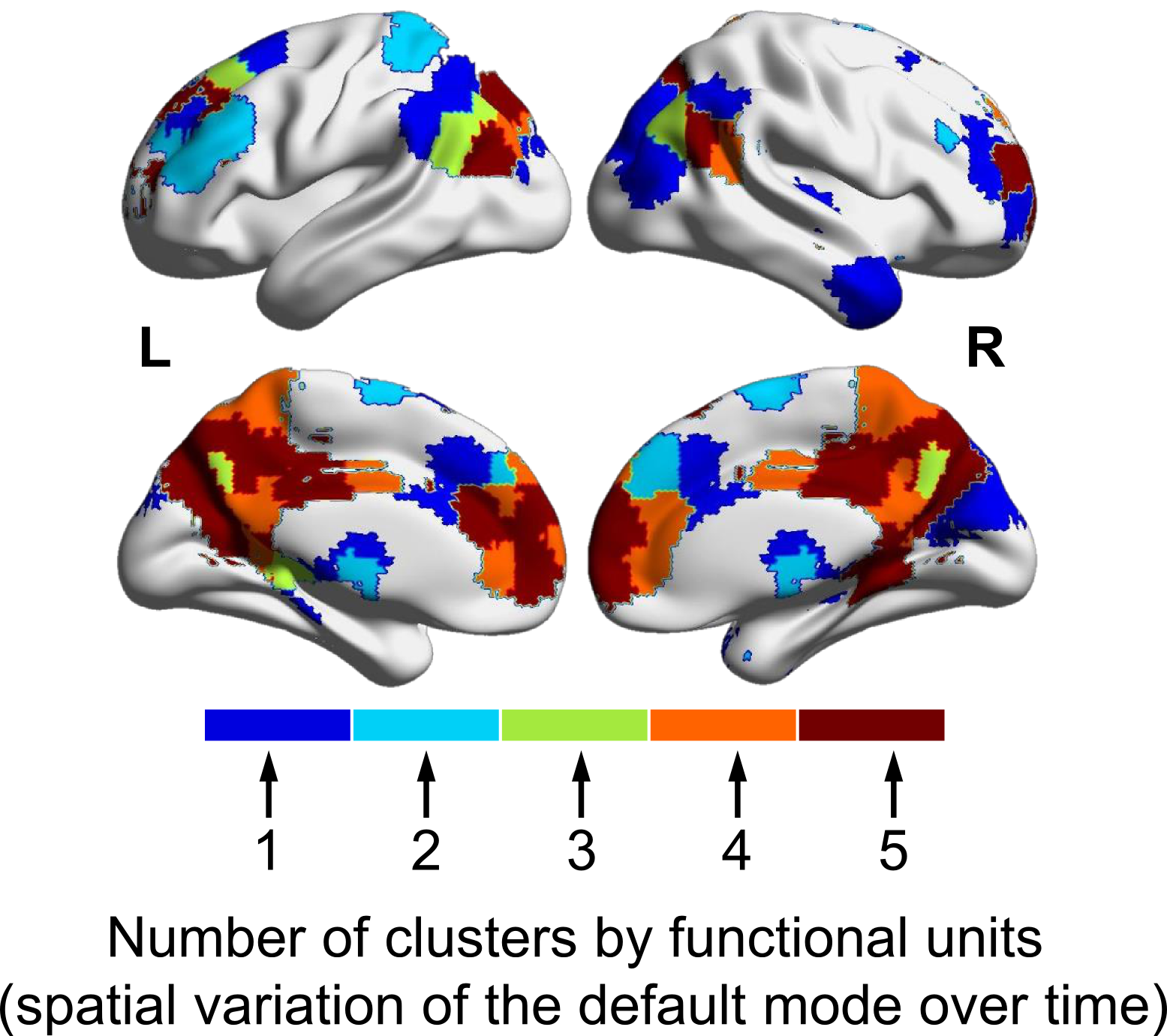
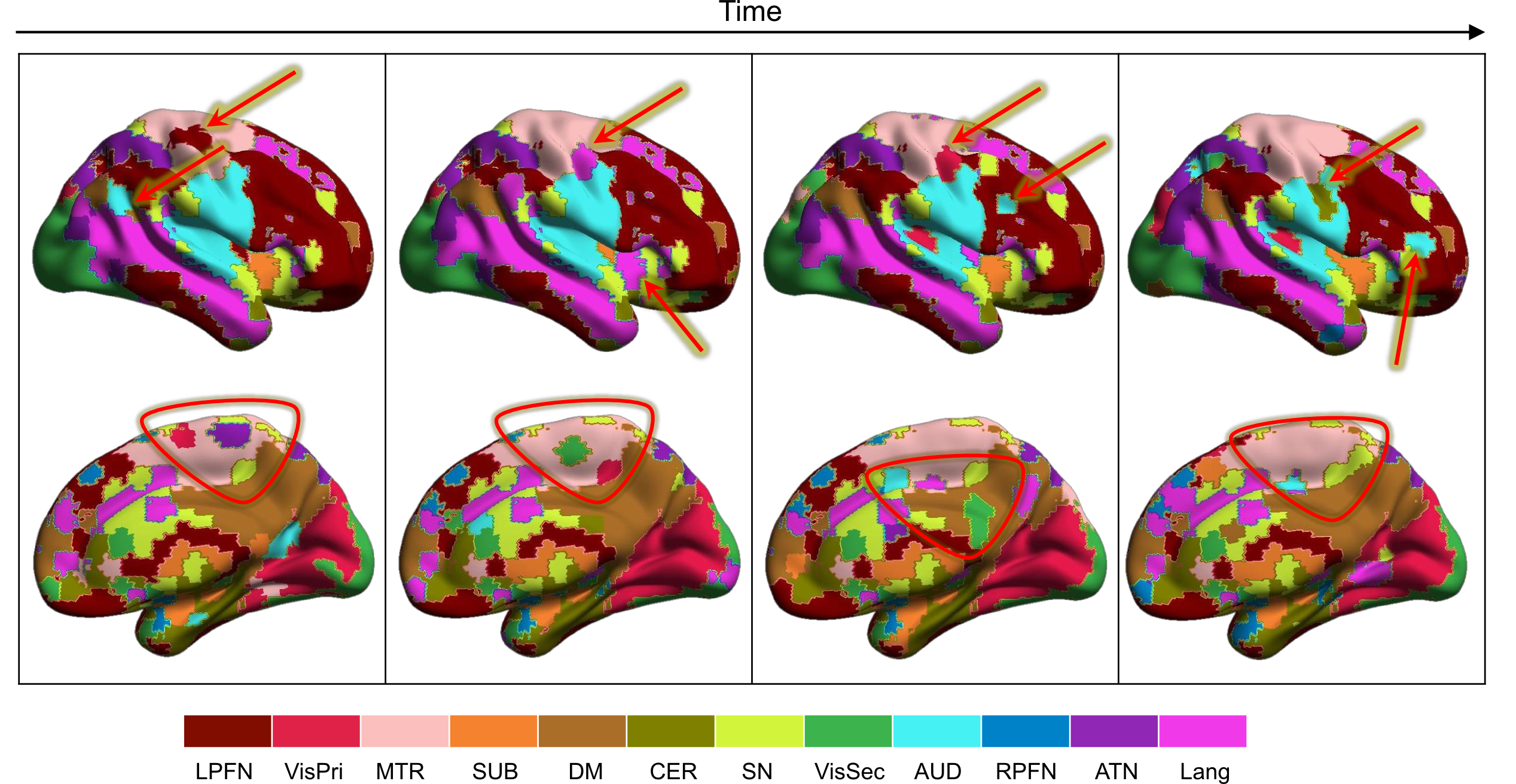
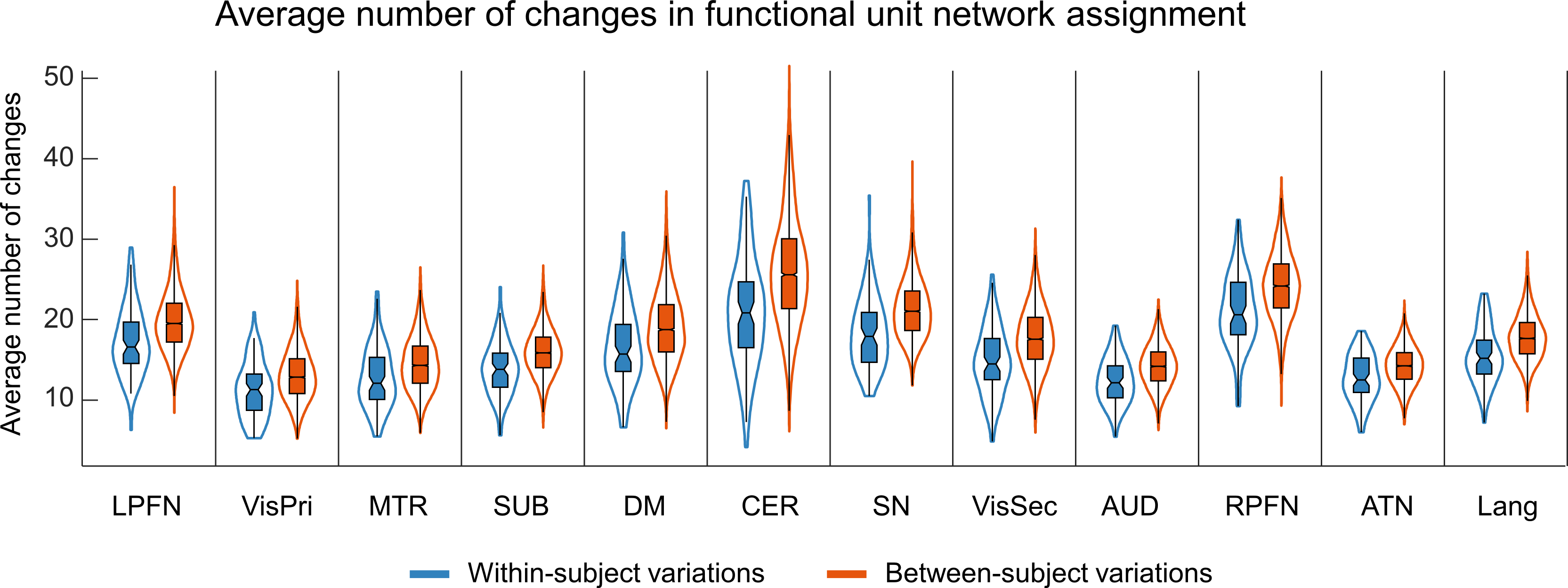
Figure 1 illustrates the distributions of FUs across the brain suggesting brain voxels can simultaneously be involved in several functional units. As the results, they could have several functional roles. Figure 2 shows an example of FUs and ability of ultra-high-order ICA to identify fine overlapping functional units. It also indicates temporal variations in FUs assignment. Interestingly, spatially adjoined FUs reveal different functional roles over time, suggesting an advantage of ICA over parcellation approaches. Further investigation shows reconfiguration in the spatial topography of brain networks over time. Figure 3 shows an example of variations in the spatial topography of brain networks. Time-windows were grouped together based on their similarity in FUs assignments to the default mode using k-medoids clustering (k = 5). The color-bar indicates the number of clusters that each region contributes to the default mode which varies from all clusters to only one cluster. Our analysis also demonstrated spatial functional reconfiguration over time across the brain (Figure 4). Spatial reconfiguration can be grouped into two categories: 1) expansion and contraction of networks spatial topography and 2) turnover in regions membership. We also compared variation in FU assignments between and within subjects. As expected, between-subjects variation is higher than within; however, the level of variations is in the same order (Figure 5). Interestingly, the results indicate high-level of agreement between variations across subjects and within subjects over time (correlation of 0.98). The network with higher (lower) variations across subjects, demonstrates lower (higher) within-subject variations. Consistent with previous findings using multi-task fMRI data, networks associated with higher cognitive functions have higher variations compare to networks involved in primary tasks.
Discussion and Conclusion
We proposed a new data-driven (atlas-free) framework that captures spatiotemporal variations of brain function at a relatively fine spatial scale and investigates the reconfiguration of functional, spatial topography both between- and within-subjects. Preliminary results demonstrate the advantage of the proposed approach in investigating variations in brain functional organizations. The findings emphasize that changes in functional connectivity go beyond sampling variability, highlighting the importance of both between and within-subject variations. Temporal reconfiguration of brain spatial topography demands further investigation of the spatial chronnectome2. Future analysis for ISMRM conference will include a) capturing spatial evolution brain functional organization over time, b) identifying the dominant brain functional topography, and c) comparing between and within-subjects spatiotemporal variations.Acknowledgements
This work was supported by grants from the National Institutes of Health grant numbers 2R01EB005846, R01REB020407, and P20GM103472; and National Science Foundation (NSF) grant 1539067 to Dr. V.D. CalhounReferences
1. Iraji, A., et al., The spatial chronnectome reveals a dynamic interplay between functional segregation and integration. bioRxiv, 2018.
2. Iraji, A., et al., Spatial dynamics within and between brain functional domains: A hierarchical approach to study time-varying brain function. bioRxiv, 2018.
Figures