0752
Sleep-related changes in diffusivity overnight: a window into glymphatic activity in humans?1University Children's Hospital, Zurich, Switzerland
Synopsis
This study examined overnight changes in diffusivity as a proxy marker for clearance in the glymphatic system, a recently proposed pathway for waste clearance in the brain. In 18 healthy adults, the mean diffusivity increased overnight in multiple brain regions, consistent with the hypothesised expansion of the extracellular space during sleep. In contrast, the diffusivity within CSF decreased overnight, possibly due to temperature effects. The overnight reduction in CSF diffusivity did not correlate with sleep parameters, but the overnight increase in diffusivity in the brain was positively correlated with the percentage of time spent in REM sleep, assessed with EEG.
Introduction
The recently proposed “glymphatic” system is thought to represent an important mechanism for clearance of waste products from the brain, whereby cerebrospinal fluid (CSF) enters the brain parenchyma via astrocytic acquaporin-4 water channels, exchanges with interstitial fluid (ISF) and flushes through the brain.1 Reports from rodent studies suggest that the sleep-wake cycle exerts a large influence on glymphatic clearance, such that the interstitial space appears to increase in volume by 60% during slow wave sleep.2
While previous MRI studies of glymphatic activity have utilized anatomical sequences in combination with intrathecal contrast agents to assess the transit of contrast through the glymphatic system,3-4 other MRI techniques like diffusion MRI are exquisitely sensitive to changes in water compartmentalization, demonstrating large signal changes in response to a change in the volume of the interstitial space. The purpose of the present study was to examine overnight changes in diffusivity as a proxy marker of glymphatic clearance, in relation to sleep parameters derived from overnight high density EEG.
Methods
The participant group consisted of 18 healthy young adult volunteers (8 female, mean age 21 years, range 18-24). On the night of the sleep EEG assessment, MR imaging investigations including diffusion tensor imaging (DTI) were performed from 8:28 pm (± 8 min), after which participants were fitted with the EEG net and sleep was recorded all night. The following morning, the MRI protocol was repeated after removal of the EEG net, at 8:12 am (± 8 min). EEG data were acquired with a high density (128 channel) EEG net (SensorNet, Electrocal Geodesic Inc., Eugene, Oregon, USA). Data were sampled at 500 Hz, referenced to the vertex electrode (Cz), and subsequently band-pass filtered (0.5-40 Hz) and downsampled to 128 Hz. Sleep stages were scored for 20-sec epochs according to standard criteria by two sleep experts.5 The duration of rapid eye movement (REM) and non-REM (slow wave) sleep was calculated for each participant as a percentage of the total sleep duration.
DTI data were collected using a pulsed gradient spin echo sequence with 35 sampling directions, using a GE 3T MR750 MRI scanner (GE Healthcare, Waukesha, WI, USA). DTI images were skull-stripped and corrected for eddy current effects, and the diffusion tensor was fitted using the tools within the FSL software library. The fractional anisotropy maps were then normalized into MNI space using the registration tools incorporated in the tbss pipeline, and the same transformations were applied to the mean diffusivity (MD) maps to align them into standard space. Differences in MD between the evening and morning scans were tested using the permutation testing methods implemented in FSL randomize, using a single group paired difference design, corrected for multiple comparisons using threshold free cluster enhancement (TFCE). The average MD within all regions showing a significant overnight change was then compared to the sleep EEG parameters using Pearson’s correlation coefficient.
Results and Discussion
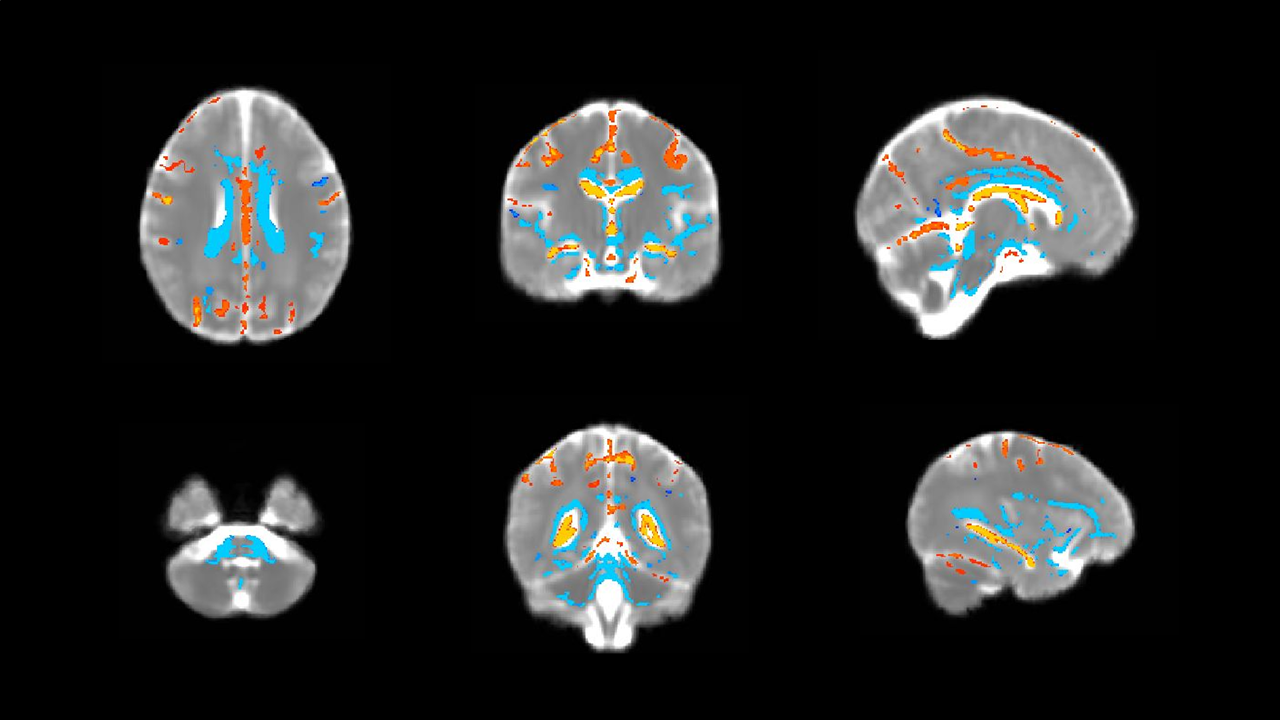
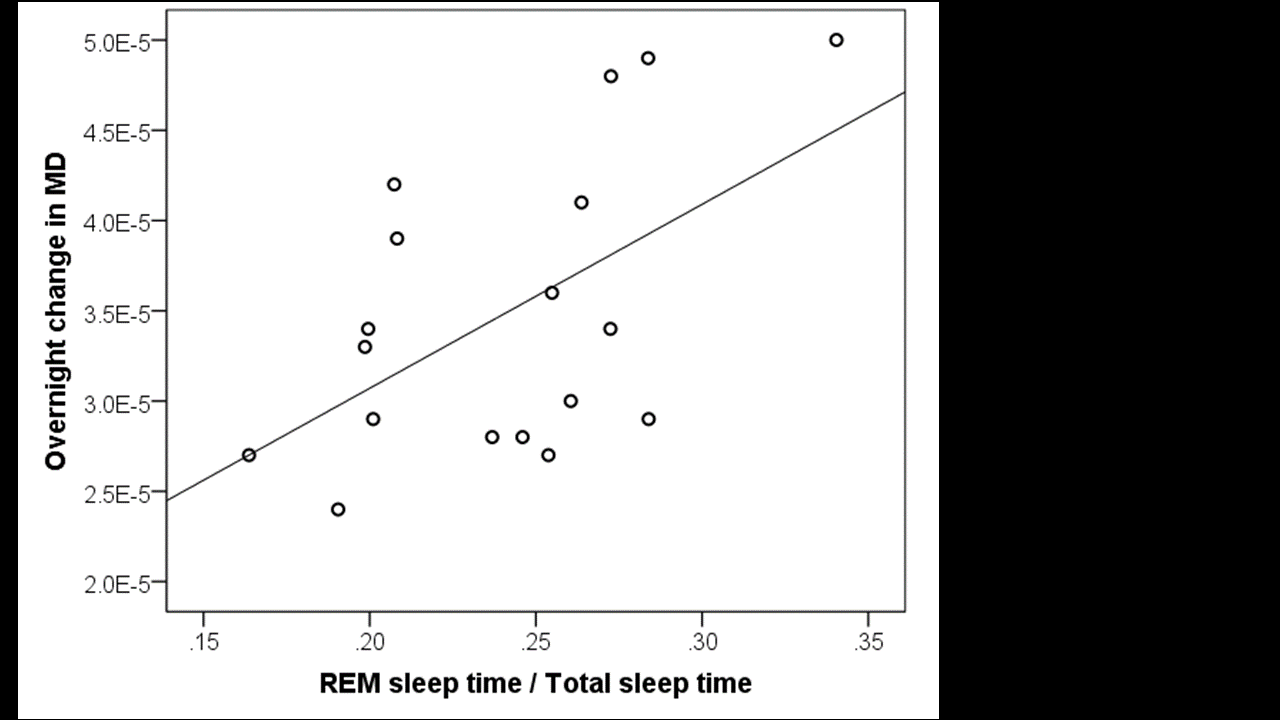
At a corrected threshold of p<0.01, MD increased overnight in regions within the cerebral white matter, basal ganglia, posterior thalamus, cerebellum, and brainstem bilaterally, consistent with an overnight expansion of the extracellular space (figure 1). Within the CSF spaces, the MD decreased overnight, possibly due to an overnight reduction in temperature, since the MD within CSF has previously been reported to represent a marker for brain temperature.6 The overnight reduction in diffusivity in the CSF spaces did not correlate significantly with any of the sleep parameters, but the overnight increase in MD in the brain was positively correlated with the percentage of time spent in REM sleep and negatively correlated with the percentage of non-REM sleep, such that participants who spent a greater fraction of sleep in a REM state showed a greater overnight increase in MD (p=0.019, figure 2). The positive correlation between the overnight change of MD and REM (rather than non-REM) sleep time is surprising in the context of reports from animal studies suggesting that that glymphatic activity increases during slow-wave sleep. However, previous rodent studies did not distinguish between REM and non-REM sleep states specifically2, and differences in glymphatic activity could also arise from inter-species differences in sleep patterns.7 Future studies will be needed to elucidate the factors underlying glymphatic clearance in humans, but the present study demonstrates that diffusion MRI combined with sleep EEG represents a promising method for evaluating glymphatic activity in the sleeping human brain.Acknowledgements
This work was supported by the Swiss National Science Foundation (320030_153387) and the Clinical Research Priority Program (CRPP) Sleep and Health of the University of Zurich.References
1. Iliff, J.J., et al., A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid beta. Sci Transl Med, 2012. 4(147): p. 147ra111.
2. Xie, L., et al., Sleep drives metabolite clearance from the adult brain. Science, 2013. 342(6156): p. 373-7.
3. Iliff, J.J., et al., Brain-wide pathway for waste clearance captured by contrast-enhanced MRI. The Journal of Clinical Investigation, 2013. 123(3): p. 1299-1309.
4. Naganawa, S., et al., Gd-based Contrast Enhancement of the Perivascular Spaces in the Basal Ganglia. Magn Reson Med Sci, 2017. 16(1): p. 61-65.
5. Feinberg, I. & Floyd, T. C. Systematic trends across the night in human sleep cycles. Psychophysiology, 1979. 16, 283-291. DOI: 10.1111/j.1469-8986.1979.tb02991.x
6. Hasan, K.M., et al., Lateral ventricular cerebrospinal fluid diffusivity as a potential neuroimaging marker of brain temperature in multiple sclerosis: a hypothesis and implications. Magn Reson Imaging, 2015. 33(3): p. 262-9.
7. Yasenkov, R. & Deboer, T. Circadian modulation of sleep in rodents. Prog Brain Res, 2012. 199:203-18.
Figures