0750
Impaired glymphatic transport and loss of peri-arterial AQP4 expression in spontaneously hypertensive stroke prone rats compared to normotensive WKY controlssunil koundal1, Simon Sanggaard1, Yuechuan Xue1, Xiaodan Liu1, Joanna Wardlaw2,3,4, Maiken Nedergaard5,6, Hedok Lee 1, and Helene Benveniste1
1Department of Anesthesiology, Yale school of medicine, new haven, CT, United States, 2Center for Clinical Brain Sciences, The University of Edinburgh, Edinburgh, United Kingdom, 3UK Dementia Research Institute, The University of Edinburgh, Edinburgh, United Kingdom, 4Row Fogo Centre for Research into Ageing and the Brain, The University of Edinburgh, Edinburgh, United Kingdom, 5Center for Translational Neuromedicine, University of Rochester Medical School, Rochester, NY, United States, 6Center for Translational Neuromedicine, University of Copenhagen, Copenhagen, Denmark
Synopsis
Cerebral small vessel disease (CSVD) is one of the important vascular factor contributing to the cognitive impairment and dementia. Clinically, CSVD hallmarks includes MR white matter hyperintensities and dilated perivascular spaces. Brain-wide perivascular transit passageways for CSF, also known as Glymphatic system has recently been described as cerebral metabolic waste clearance pathway. We evaluated Glymphatic transport by DCE-MRI in middle aged spontaneously hypertensive stroke prone (SHRSP) rats and normal Wistar Kyoto (WKY) rats, which demonstrated significant impairment of Glymphatic transport in brain parenchyma in 7-month old SHRSP rats in comparison to controls.
Introduction
Cerebral small vessel disease (SVD) is now recognized as one of the most important vascular contributors to cognitive impairment and dementia (VCID) (1,2). The cause of VCID from cerebral SVD is still incompletely understood, however untreated hypertension is considered a major risk factor (3). Several MRI hallmarks are associated with clinical SVD diagnosis including white matter hyperintensities and dilated perivascular spaces (PVS) (4). On the basis of these MRI observations several mechanisms including breakdown of the blood brain barrier, perivascular inflammation and dysfunction of the glymphatic pathway have been suggested as mechanisms that trigger SVD (1). The glymphatic system is a brain-wide perivascular transit passageway for cerebrospinal fluid (CSF) which facilitates waste drainage in a manner driven by aquaporin-4 water channels (AQP4) (5). The AQP4 channels, strategically positioned perivascularly, on astrocytic end-feet, help propel CSF into the interstitial fluid (ISF) thereby facilitating CSF-ISF mixing and waste drainage. Hypothetically, untreated hypertension with stiffening of small arterioles, perivascular inflammation and AQP4 channel demise could impair glymphatic metabolic waste drainage thereby exacerbating perivascular fluid accumulation as is observed in SVD in form of dilated PVS. Here we test this hypothesis in untreated spontaneously hypertensive stroke prone (SHRSP) rats and normal Wistar Kyoto (WKY) rats. Further, we hypothesize that glymphatic transport is compromised in SHRSP rats, secondary to decreased polarization of perivascular AQP4 water channels.Methods
All animal experiments were approved by the Yale University IACUC. Seven-month old SHRSP (N=7) and WKY (N=6) female rats (Charles Rivers, Mass) were used for the experiments. Under surgical plane anesthesia, a small catheter was implanted into the CSF via the cisterna magna (6). Images were acquired on a Bruker 9.4T/16 MRI equipped with a volume transmit/receive or surface receive only coils interfaced with Paravision 6. All rats were scanned while anesthetized with dexmedetomidine supplemented with low-dose isoflurane and physiological parameters continuously monitored (7). The dynamic contrast-enhanced MRI (DCE-MRI) during and after the infusion of gadoteric acid (Gd-DOTA) was applied to produce dynamic measurement of glymphatic transport (8). Following MRI, the rats were perfusion fixed with formalin and the brains were examined for GFAP, AQP4, and aSMA expression using immunofluorescence staining. To calculate distribution volume (VT) of Gd-DOTA representing glymphatic transport (8), the time-signal curves (TSCs) from the CSF (representing the main “input” function to the brain) and from the whole brain were extracted from each rat and the Logan plot (9) was applied using kinetic modeling PMOD software (version 3.9).Results
The mean arterial blood pressure in unanesthetized SHRSP rats at 7 months was ≈120-140mmHg, while WKY was in normal range ~90-100mmHg. The average TSCs of CSF-Gd (% signal change from baseline) extracted from the basal cisterns from WKY rats were similar to that of SHRSP rats indicating that CSF transport in this compartment is comparable between the two strains (results not shown). Fig. 1A shows the average TSCs extracted from brain over the 2.5 hr experimental time window demonstrating that the Gd-enhanced brain parenchymal signal amplitude is significantly reduced in SHRSP rats compared to WKY indicating lower glymphatic transport capabilities in the hypertensive rats. Glymphatic transport as assessed by the Gd-DOTA tissue distribution volume (VT) calculated by the Logan plot demonstrated almost 2-fold lower glymphatic transport in SHRSP rats compared to the WKY rats (VT WKY (N=7) = 0.230 ± 0.023 versus VT SHRSP (N=6) = 0.133 ± 0.016, P <0.005). Fig. 1B shows 3D color-coded, volume rendered maps of Gd uptake in a WKY and SHRSP rats at 60 min and clearly shows the more efficient transport in the non-hypertensive WKY compared to the hypertensive SHRSP rat. Fig. 2 shows the AQP4 immunoreactivity in a WKY (2A) rat compared to SHRSP (2B rat at the level of the dorsal hippocampus demonstrating that the AQP4 expression pattern is different between the two strains. Specifically, in the SHRSP rats AQP4 expression around the penetrating arterioles were ‘dispersed’ (i.e. lost their perivascular polarity) similarly to what has been described in aging rodent brain (10).Discussion & Conclusions
We evaluated glymphatic transport by DCE-MRI in middle aged SHRSP and WKY rats and demonstrated significant impairment of glymphatic transport in brain parenchyma in 7-month old SHRSP rats when compared to controls. Major drivers of glymphatic transport include vascular pulsatility(11), state of arousal(12), and AQP4 water channel polarization to astrocytic end-feet(5). In our experiments, SHRSP as well as WKY rats were anesthetized with the same anesthetic regimen and physiological parameters were within similar ranges. Hence, impaired glymphatic transport observed in SHRSP rats is most likely due to loss of peri-arterial AQP4 polarization secondary to chronic hypertension and remodeling of the perivascular space.Acknowledgements
The present work was supported by National Institutes of Health RF-AG053991, RF-AG057705, R01-NS100366, and Foundation Leducq Transatlantic Network of Excellence (16/CVD/05).References
1. Brown R, Benveniste H, Black SE, et al. Understanding the role of the perivascular space in cerebral small vessel disease. Cardiovasc Res. 2018. 2. Ter Telgte A, van Leijsen EMC, Wiegertjes K, Klijn CJM, Tuladhar AM, de Leeuw FE. Cerebral small vessel disease: from a focal to a global perspective. Nat Rev Neurol. 2018;14:387-398. 3. van Middelaar T, Argillander TE, Schreuder F, Deinum J, Richard E, Klijn CJM. Effect of Antihypertensive Medication on Cerebral Small Vessel Disease: A Systematic Review and Meta-Analysis. Stroke. 2018;49:1531-1533. 4. Wardlaw JM, Smith EE, Biessels GJ, et al. Neuroimaging standards for research into small vessel disease and its contribution to ageing and neurodegeneration. Lancet Neurol. 2013;12:822-838. 5. Iliff JJ, Wang M, Liao Y, et al. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid beta. Sci Transl Med. 2012;4:147ra111. 6. Lee H, Mortensen K, Sanggaard S, et al. Quantitative Gd-DOTA uptake from cerebrospinal fluid into rat brain using 3D VFA-SPGR at 9.4T. Magn Reson Med. 2018;79:1568-1578. 7. Iliff JJ, Lee H, Yu M, et al. Brain-wide pathway for waste clearance captured by contrast-enhanced MRI. J Clin Invest. 2013;123:1299-1309. 8. Lee H, Xie L, Yu M, et al. The Effect of Body Posture on Brain Glymphatic Transport. J Neurosci. 2015;35:11034-11044. 9. Logan J, Fowler JS, Volkow ND, et al. Graphical analysis of reversible radioligand binding from time-activity measurements applied to [N-11C-methyl]-(-)-cocaine PET studies in human subjects. J Cereb Blood Flow Metab. 1990;10:740-747. 10. Kress BT, Iliff JJ, Xia M, et al. Impairment of paravascular clearance pathways in the aging brain. Ann Neurol. 2014;76:845-861. 11. Iliff JJ, Wang M, Zeppenfeld DM, et al. Cerebral arterial pulsation drives paravascular CSF-interstitial fluid exchange in the murine brain. J Neurosci. 2013;33:18190-18199. 12. Xie L, Kang H, Xu Q, et al. Sleep drives metabolite clearance from the adult brain. Science. 2013;342:373-377.Figures
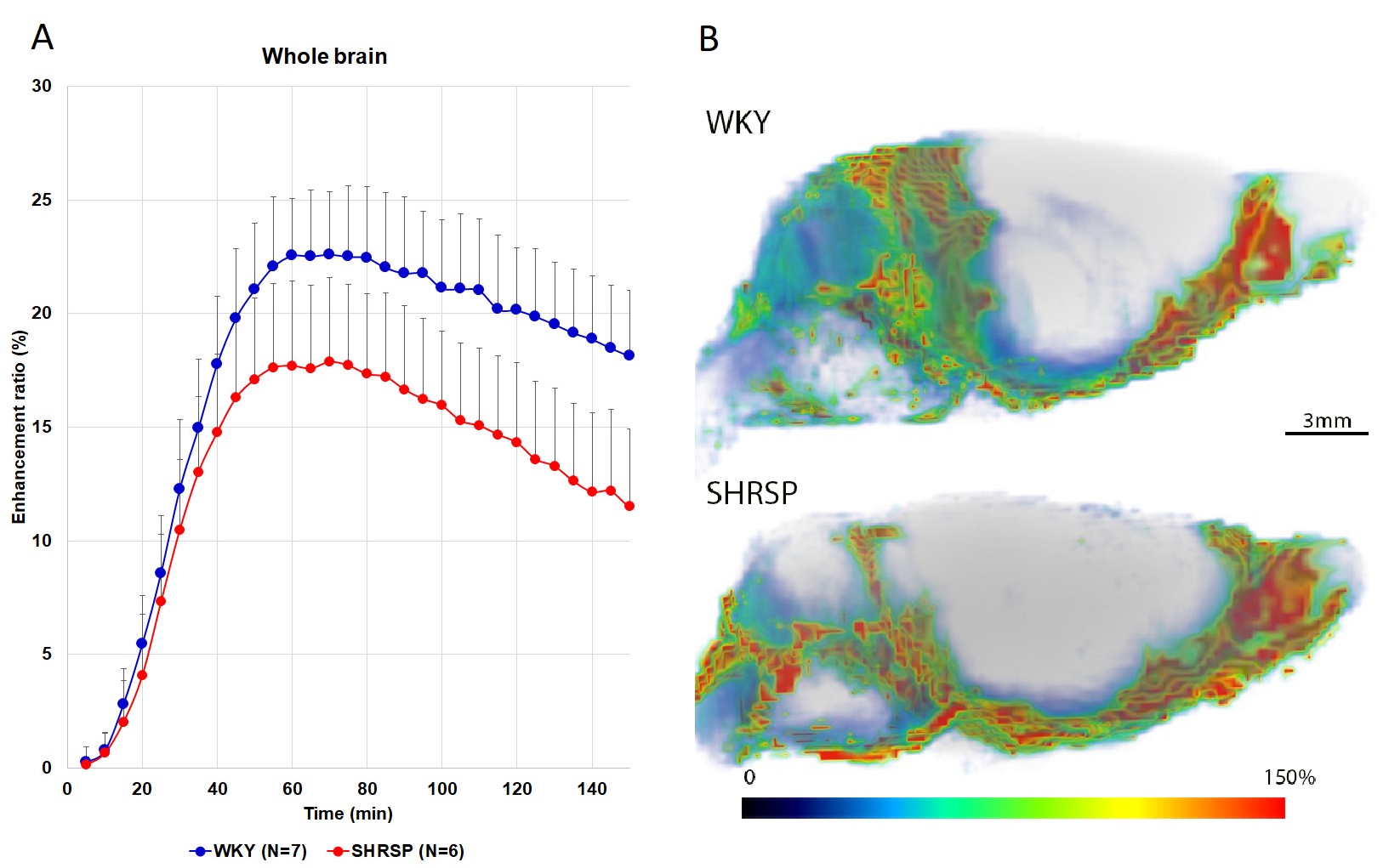
Fig.
1A: Average TSCs extracted from whole brain demonstrating
that the Gd-enhanced brain parenchymal signal amplitude is significantly
reduced in SHRSP rats compared to WKY indicating lower glymphatic transport
capabilities in the hypertensive rats. Data are mean ± SD. Fig.
1B: 3D color-coded, volume rendered maps of Gd uptake in a
WKY and SHRSP rats at 60 min demonstrating more efficient Gd transport into
brain parenchyma of the WKY compared to the hypertensive SHRSP rat. The
color-coded scale represents the % enhancement ratio.
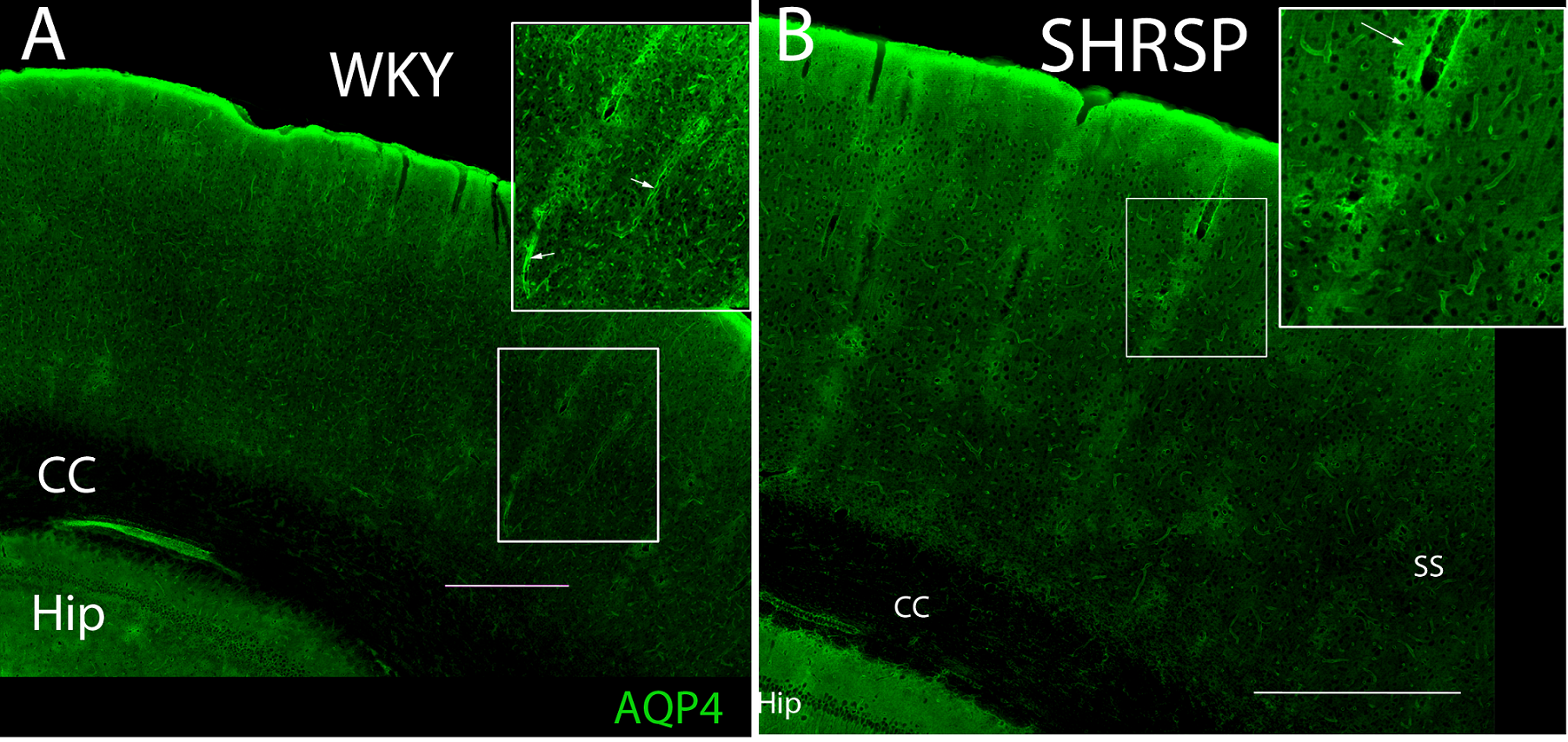
Fig. 2A: AQP4 immuno-activity in a WKY rat at the level of the dorsal hippocampus. AQP4 is associated with all vessels and there is high AQP4 expression perivascularly. Fig. 2B: AQP4 immuno-activity in a SHRSP rat at the level of the dorsal hippocampus. The AQP4 expression pattern in the somatosensory cortex (SS) is different compared to the WKY rat. Specifically, AQP4 expression along the penetrating arterioles is less polarized and more dispersed. CC=corpus callosum, Hip=hippocampus, SS=somatosensory cortex. Scale bar = 500 micron