0719
Hyperpolarized [1-13C] Alanine Ethyl Ester for Assessment of Hepatic Alanine Metabolism1Advanced Imaging Research Center, UT Southwestern Medial Center at Dallas, Dallas, TX, United States, 2Radiology, UT Southwestern Medical Center at Dallas, Dallas, TX, United States, 3Electrical Engineering, University of Texas at Dallas, Dallas, TX, United States
Synopsis
[1-13C] alanine ethyl ester was studied as hyperpolarized substrate to measure the alanine metabolism in rat liver. The results show that [1-13C] alanine ethyl ester enters the cell converts to [1-13C] lactate more efficiently. Therefore, [1-13C] alanine ethyl ester is a potential compound to assess hepatic alanine metabolism with improved sensitivity.
Introduction
Alanine
is the principal amino acid released by skeletal muscle and taken up by the
liver. It is the most important amino
acid donor to hepatic gluconeogenesis. Liver accumulates plasma alanine and
converts it to pyruvate, which is oxidized to
lactate and/or enters gluconeogenesis pathway. Liver is the major site
of gluconeogenesis and contains considerably high titer of gluconeogenic
enzymes and low glycolytic enzymes [1]. Tracing alanine metabolism in vivo can provide insights into metabolic
changes in liver complications including hepatitis infection,
fibrosis, hepatocellular carcinoma. Hyperpolarized 13C NMR can
provide a rapid assessment of metabolic enzyme activities in vivo [2]. Previous studies of hyperpolarized [1-13C]
alanine demonstrated its usefulness to investigate hepatic alanine metabolism [3, 4]. In particular, [1-13C] lactate
produced from hyperpolarized [1-13C] alanine in liver was primarily
from intracellular hepatic metabolism, allowing in vivo assessment of intracellular redox-state from the ratio of
[1-13C] lactate to [1-13C] pyruvate. However
intracellular concentration of [1-13C] alanine
is limited by the activity of alanine-serine-cysteine transporter [5]. In this study, we developed hyperpolarized [1-13C]
alanine
ethyl ester as a more potent agent for investigating liver metabolism because
it is hypothesized to cross biological membranes more readily than alanine and,
thus, to increase MR sensitivities of the product.
Methods
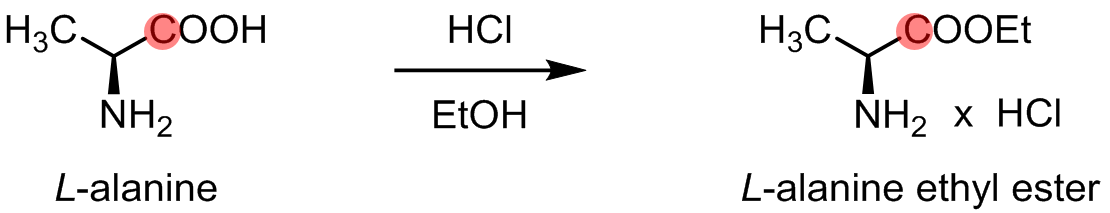
[1-13C] alanine ethyl ester was synthesized by esterification of commercially available (Sigma Aldrich) [1-13C] L-alanine (Fig. 1). The synthesis and the chemical shift of [1-13C] alanine ethyl ester were confirmed by 13C NMR at 9.4T. A GE SPINlab polarizer was used for dynamic nuclear polarization of [1-13C] alanine and [1-13C] alanine ethyl ester. Both in vitro polarization measurements and in vivo animal MR spectroscopy were performed at a clinical 3T MR scanner (GE Discovery 750W). 6.2M of [1-13C] alanine ethyl ester was prepared in 3:1 w/w water : glycerol with 15mM OX063. The dissolved sample was adjusted to pH 7.5 with 500ul of 250mM Tris-HCl buffer. The liquid-state polarization levels and longitudinal relaxation times (T1) of the hyperpolarized substrates were estimated as previously described [6]. For the in vivo studies, healthy male Sprague-Dawley rats (200-400g) were used. A custom-built 13C surface coil (single loop, Ø = 28mm) was placed on top of the liver area for both radiofrequency (RF) and data acquisition. 80-mM hyperpolarized [1-13C] alanine or [1-13C] alanine ethyl ester was injected intravenously as a bolus (1 mmol/kg body weight, up to 4.0 mL, injection rate = 0.25 mL/s), immediately followed by a dynamic 13C MRS scan (FID CSI, 10o hard pulse RF excitation, repetition time = 3s, scan time = 4min).Results and Discussion
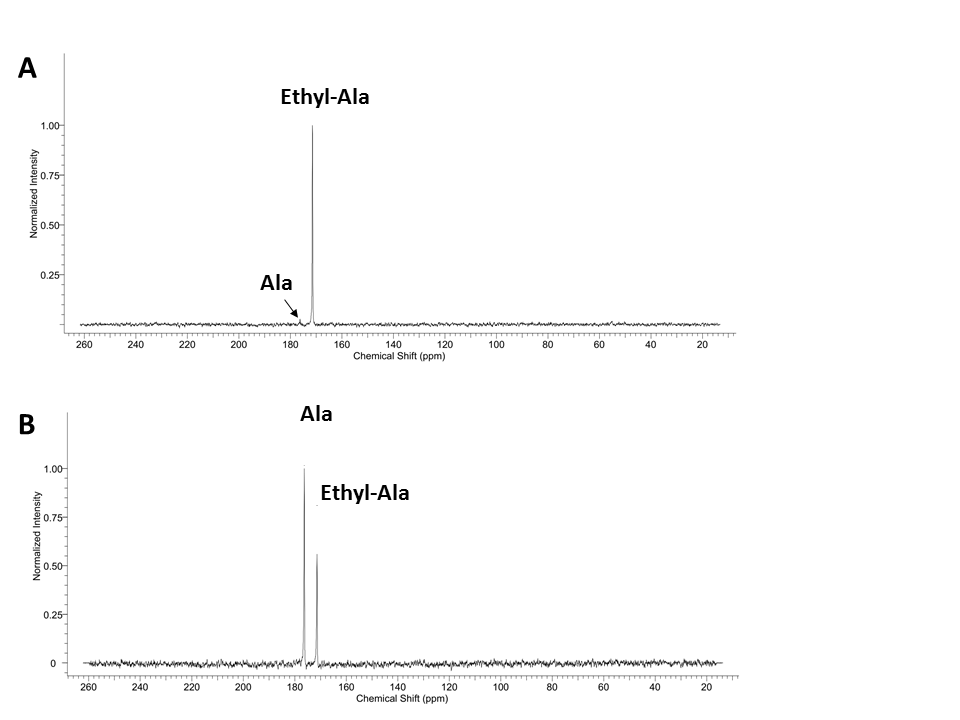
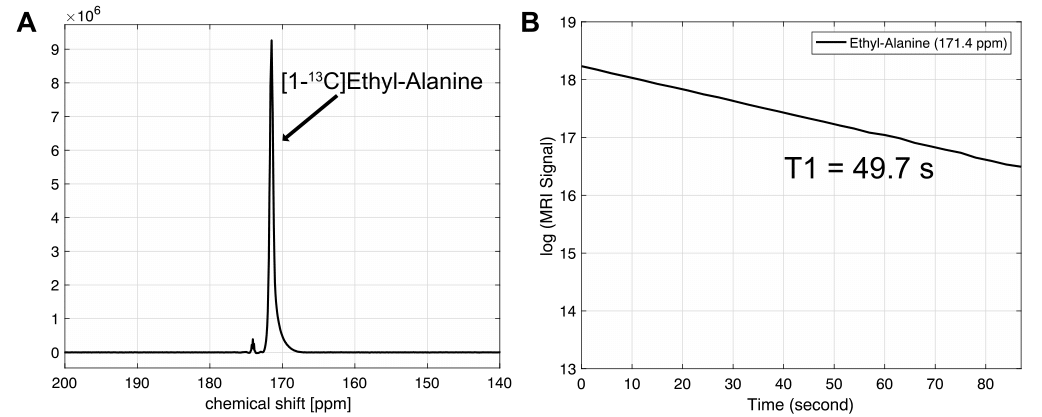
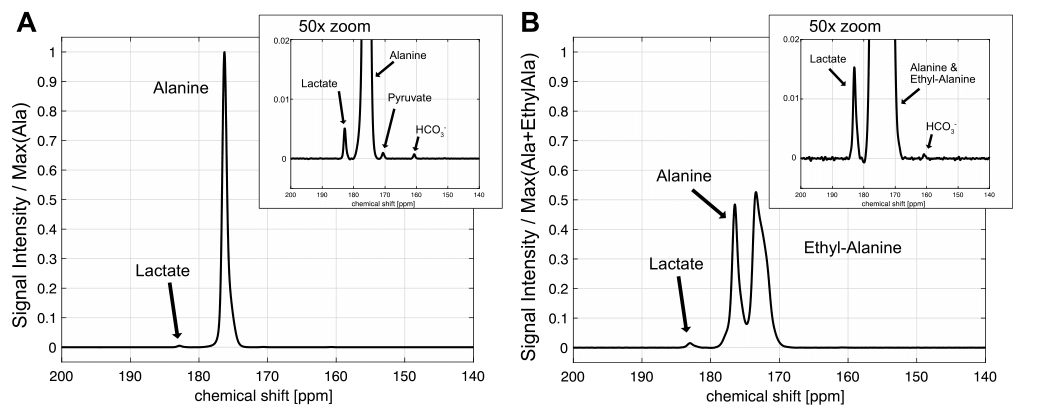
Fig. 2 shows the chemical shifts of the synthesized [1-13C] alanine ethyl ester and small residual of [1-13C] alanine. The chemical shift of [1-13C] alanine ethyl ester (171.4 ppm) was 4.9 ppm upfield compared to the [1-13C] alanine resonance (176.3 ppm). In order to validate whether the additional metabolite is [1-13C] alanine, [1-13C] alanine was added to the [1-13C] alanine ethyl ester sample as standard (Fig. 2B). The liquid-state polarization level at the time of dissolution was estimated as 6.9-17.3 % and the T1 of [1-13C] alanine ethyl ester was measured as ~50 s (Fig. 3). Fig. 4 shows the time-averaged 13C spectra in rat liver acquired after an injection of hyperpolarized [1-13C] alanine (A) or [1-13C] alanine ethyl ester (B). Significantly larger [1-13C] lactate peak was detected from hyperpolarized [1-13C] alanine ethyl ester than hyperpolarized [1-13C] alanine (more than two-fold). [1-13C] pyruvate production could not be detected as the peak was overlapped with [1-13C] alanine ethyl ester peak. Bicarbonate detection was inconsistent depending on the nutritional state of the animals and the polarization level. Unlike the in vitro studies (Fig. 3),alanine ethyl ester was rapidly hydrolyzed into alanine in vivo, generating a large peak of [1-13C] alanine (Fig. 4B). This can be due to the rapid de-esterification of alanine ethyl ester while crossing the cell membrane or in the blood. Indeed, rodents have high amounts of plasma carboxylesterase (EC 3.1.1.1). We expect to have liver-specific alanine signals and more sensitive lactate (and bicarbonate) measurements as human plasma does not contain carboxylesterase [7, 8]. The future study will focus on optimizing alanine ethyl ester sample for hyperpolarization and identifying the de-esterification mechanism of alanine ethyl ester in blood.Conclusion
Significantly larger 13C-lactate was produced from hyperpolarized [1-13C] alanine ethyl ester than hyperpolarized [1-13C] alanine, indicating that the ethyl ester form of alanine indeed crosses the plasma membrane and enter the gluconeogenesis cycle more efficiently than the original form of the amino acid. The results suggest that [1-13C] alanine ethyl ester is a more sensitive probe to trace the liver metabolism non-invasively under pathophysiological conditions and help diagnosis or monitor disease progression.Acknowledgements
Funding: National Institutes of Health of the United States (P41 EB015908, S10 OD018468); The Mobility Foundation; The Texas Institute of Brain Injury and Repair.References
1. Wisniewski, J.R., A. Gizak, and D. Rakus, Integrating Proteomics and Enzyme Kinetics Reveals Tissue-Specific Types of the Glycolytic and Gluconeogenic Pathways. J Proteome Res, 2015. 14(8): p. 3263-73.
2. Kurhanewicz, J., et al., Analysis of cancer metabolism by imaging hyperpolarized nuclei: prospects for translation to clinical research. Neoplasia, 2011. 13(2): p. 81-97.
3. Park, J.M., et al., In vivo assessment of intracellular redox state in rat liver using hyperpolarized [1-(13) C]Alanine. Magn Reson Med, 2017. 77(5): p. 1741-1748.
4. Hu, S., et al., In vivo measurement of normal rat intracellular pyruvate and lactate levels after injection of hyperpolarized [1-(13)C]alanine. Magn Reson Imaging, 2011. 29(8): p. 1035-40.
5. Joseph, S.K., N.M. Bradford, and J.D. McGivan, Characteristics of the transport of alanine, serine and glutamine across the plasma membrane of isolated rat liver cells. Biochem J, 1978. 176(3): p. 827-36.
6. Mayer, D., et al., Application of hyperpolarized [1-(1)(3)C]lactate for the in vivo investigation of cardiac metabolism. NMR Biomed, 2012. 25(10): p. 1119-24.
7. Li, B., et al., Butyrylcholinesterase, paraoxonase, and albumin esterase, but not carboxylesterase, are present in human plasma. Biochem Pharmacol, 2005. 70(11): p. 1673-84.
8. Yang, D., et al., Human carboxylesterases HCE1 and HCE2: ontogenic expression, inter-individual variability and differential hydrolysis of oseltamivir, aspirin, deltamethrin and permethrin. Biochem Pharmacol, 2009. 77(2): p. 238-47.
Figures