0614
De-noising high-resolution fMRI data using cortical depth-dependent analysis1Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts General Hospital, Boston, MA, United States, 2Department of Radiology, Harvard Medical School, Boston, MA, United States, 3Harvard-Massachusetts Institute of Technology Division of Health Sciences and Technology, Cambridge, MA, United States, 4Biomedical Engineering, Boston University, Boston, MA, United States
Synopsis
In this study, we exploit cortical depth-dependent information to de-noise high-resolution fMRI data. We have proposed a novel de-noising pipeline that can automatically differentiate neural and non-neural fluctuations according to cross-cortical-depth delay patterns. We also show that by excluding voxels intersecting the pial surface can reduce physiological effects and improve neuronal/spatial specificity.
Introduction
High-resolution fMRI at ultra-high fields (≥7T) enables the investigation of functional architecture at fine spatial scales, and permits new strategies for improving sensitivity and neuronal specificity through the removal of physiological artifacts and noise. Here we propose to differentiate neural and non-neural fluctuations according to cross-cortical-depth delay patterns, and demonstrate its efficacy in de-noising visual task data. We also show that restricting analyses to voxels deeper in the cortex can mitigate systemic physiological fluctuations, and may potentially improve the spatial specificity of fMRI results.Methods
1. Using cross-cortical-depth delays to differentiate neural and non-neural fluctuations
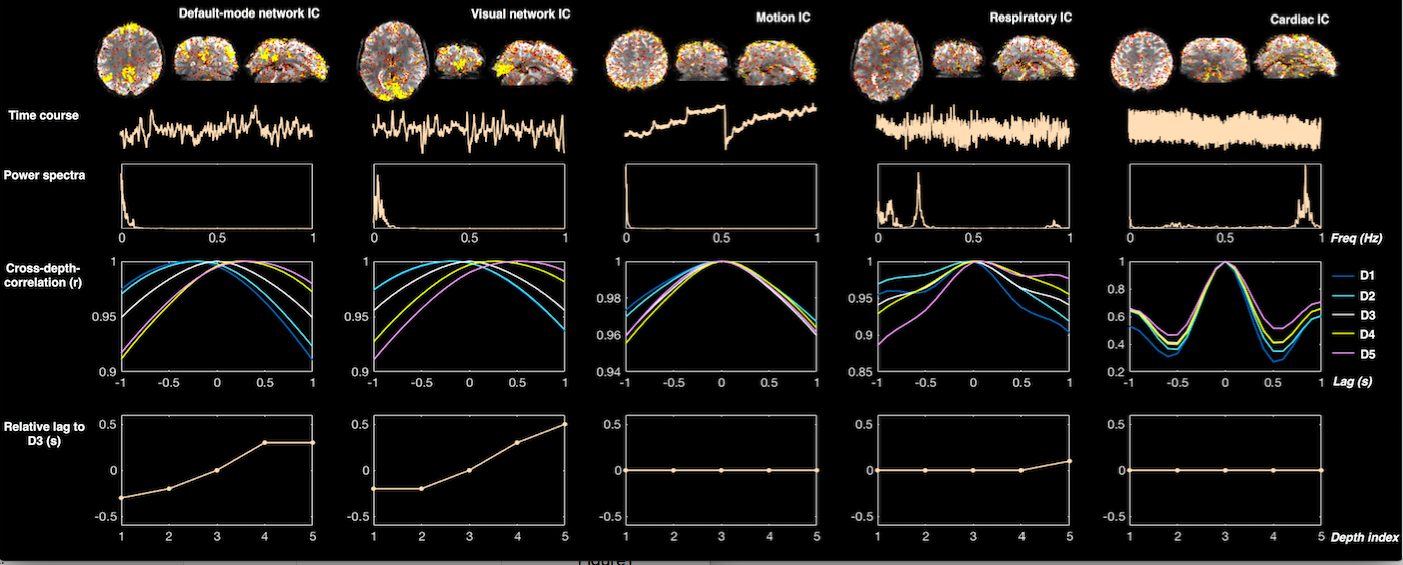
Rationale: Because neural signals initiate within the parenchyma, and the BOLD signal propagates downstream along draining veins toward the pial surface[1,2], BOLD time course from superficial cortical layers should therefore lag deeper layers. In contrast, such a temporal progression may not exist in non-neural fluctuations.
Data: Four healthy subjects (3F/1M, 28±6 y.o.) provided informed consent and were imaged on a whole-body 7T scanner (Siemens Healthineers, Erlangen, Germany) with a custom-built 32-channel receive coil array. A 0.75 mm isotropic FOCI-MEMPRAGE[3] was acquired for cortical surface reconstruction using Freesurfer. Each subject underwent three visual task scans (4 task blocks, 16/24 s on/off per block) with graded spatial and temporal resolutions: voxel size=1.5/2.0/3.0 mm iso., TR=1500/1040/745 ms, FA=68/60/52o, TE=26 ms, SMS factor=3.
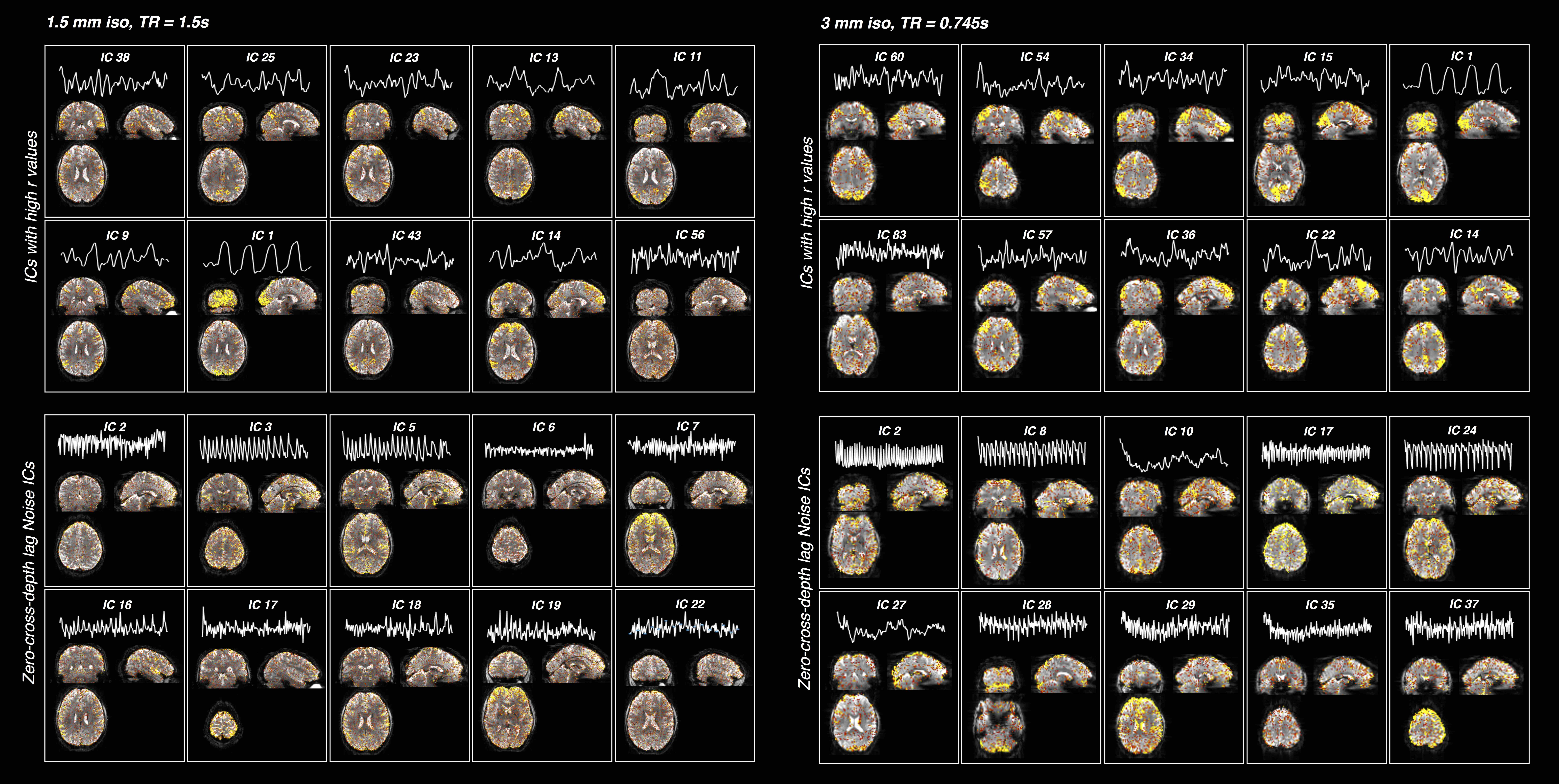
Analysis: Surface-based cortical depth estimation was performed with the high-resolution T1 data[3], following which the normalized cortical depth (‘0’:white matter; ‘1’:pial) of each EPI voxel was computed according to its centroid coordinate. Spatial decomposition using ICA: after rigid-body realignment and removal of scan drifts, each functional scan was spatially decomposed using MELODIC ICA, with the optimal component number automatically chosen[4]. Identifying nuisance (non-neural) ICs according to cross-cortical-depth delay patterns: for each IC, voxels within each network were separated into five groups based on their normalized cortical depths (D1:0–40%, D2:40–80%, D3:80–120%, D4:120–160%, D5:160–200%, where depths >100% denote locations above the pial). Relative delays across the mean signals of each cortical depth were then estimated using temporal cross-correlation. Finally, to quantify whether superficial cortical layer lags deeper layer, the relative delay vector from depths D1–D5 was correlated with an order vector (1:5) using Spearman’s correlation coefficient r, and ICs with low values of r were considered non-neural. Visual task activation: nuisance ICs (r<0.2) were included along with task regressors in a general linear model (GLM) to infer task activation, with noise autocorrelations estimated by SPM’s FAST option[5].
2. Resolving purely-cortical activity to reduce systemic physiological noise and improve spatial specificity
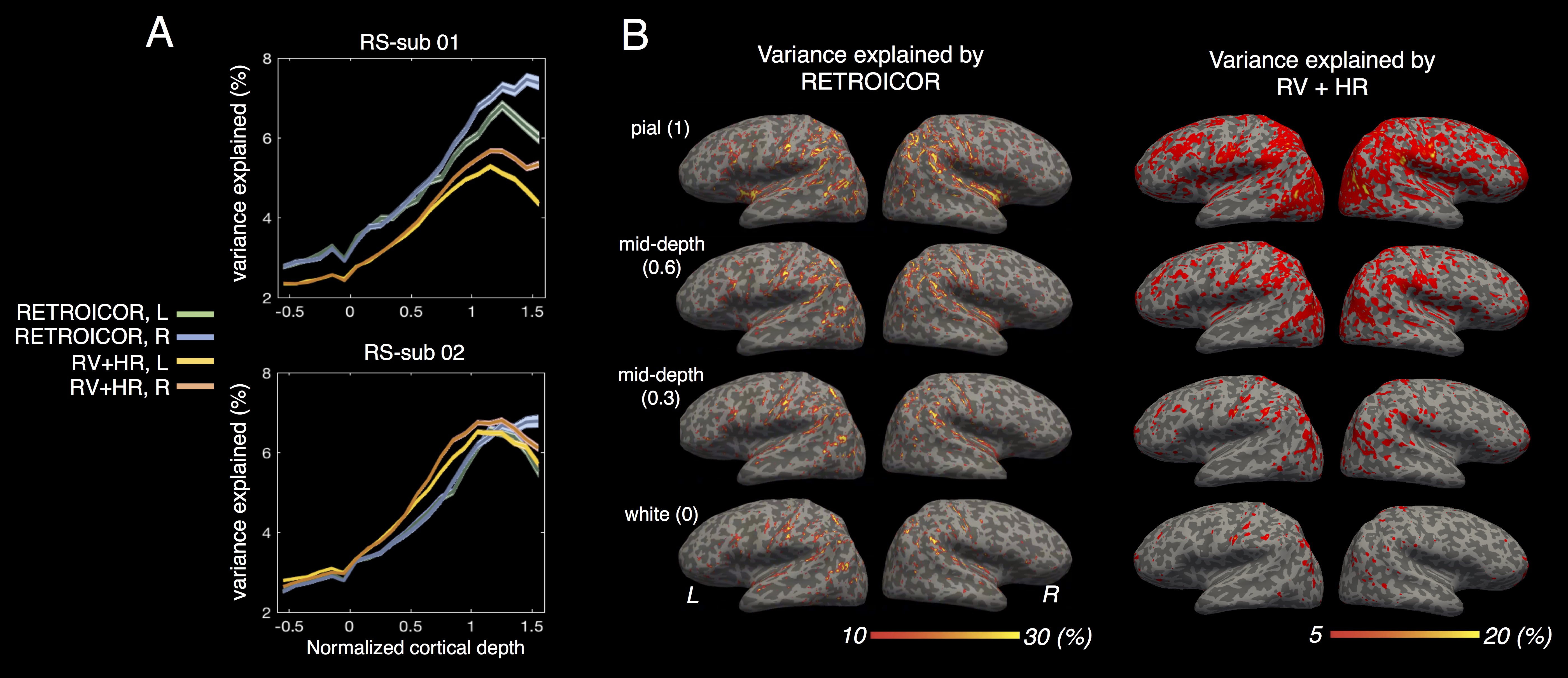
Two healthy subjects each underwent a 10-min resting-state scan (1.5 mm iso., TR/TE=1500/26 ms). Quantifying fractional contributions of physiological noise across depths: quasi-periodic respiratory and cardiac fluctuations, and slow dynamics triggered by respiratory volume (RV) and heart rate (HR) changes, were modelled using RETROICOR[6] and RVHRCOR[7], then regressed against each voxel’s time series. Resting-state networks (RSNs) at deep depths were resolved using cortex-based “masked-ICA”[8], then projected onto the cortical surface at different depths for visualization.
Results & Discussion
Fig. 1 illustrates the distinct cross-cortical-depth delay patterns of neural and non-neural fluctuations. Fig. 2 shows ICs with the highest r values (“relevant ICs”) and with zero cross-cortical-depth lags (“nuisance ICs”) in one exemplar subject. At both spatial resolutions (1.5 mm and 3 mm), “relevant ICs” generally exhibit structured spatial patterns that resemble networks whereas “nuisance ICs” associate more with physiological waveforms or well-characterized motion-like spatial patterns.
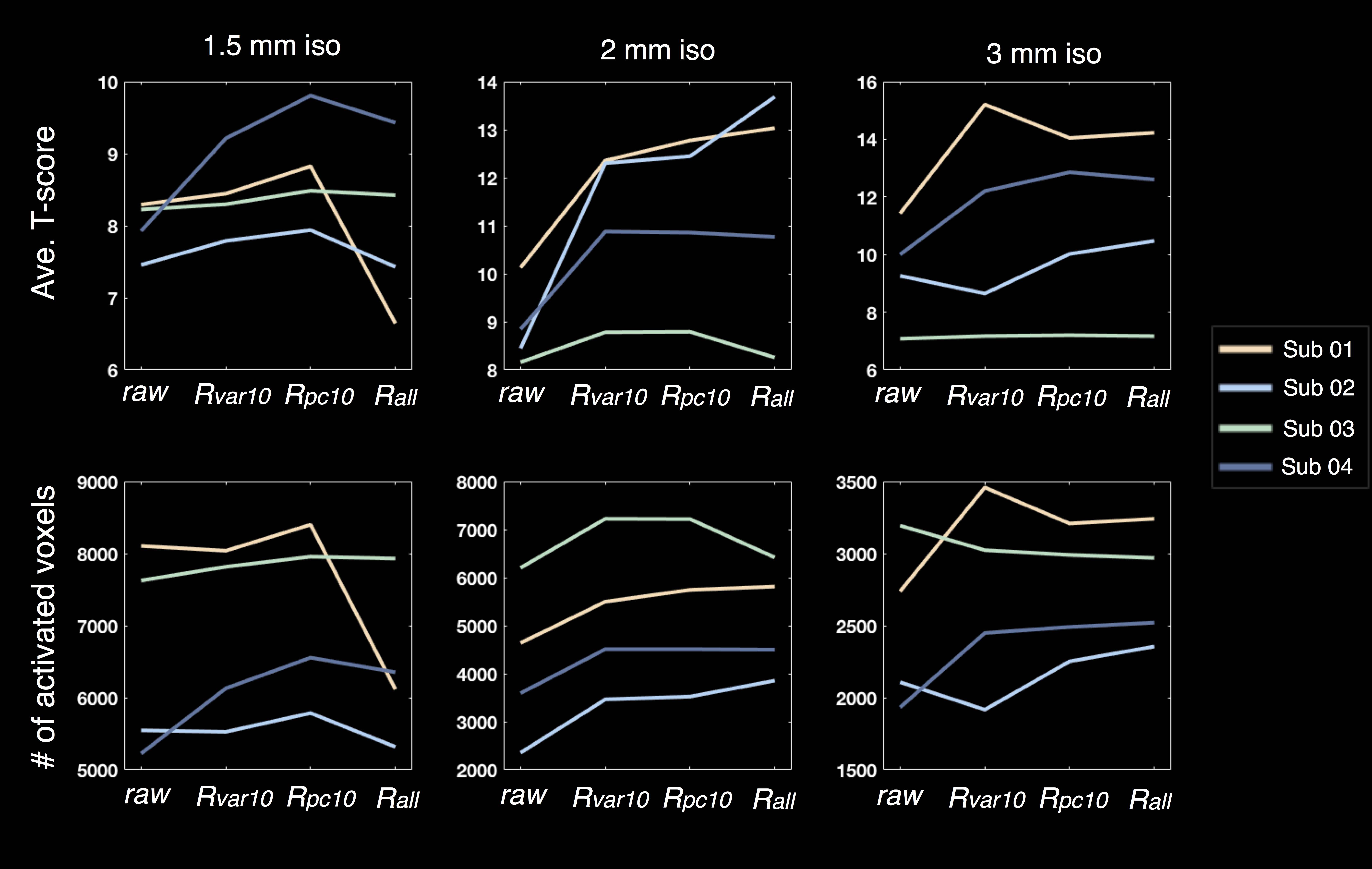
A notable trend of enhanced T-scores in task activation is achieved when including nuisance ICs in GLM (Fig. 3 ‘raw’ vs. ‘Rall’), suggesting the efficacy of the proposed de-noising approach. For data with fewer temporal points, restricting nuisance regressors to most pivotal ones is preferable (Fig. 3, ‘Rvar10’ and ‘Rpc10’ vs. ‘raw’ at 1.5 mm iso.).
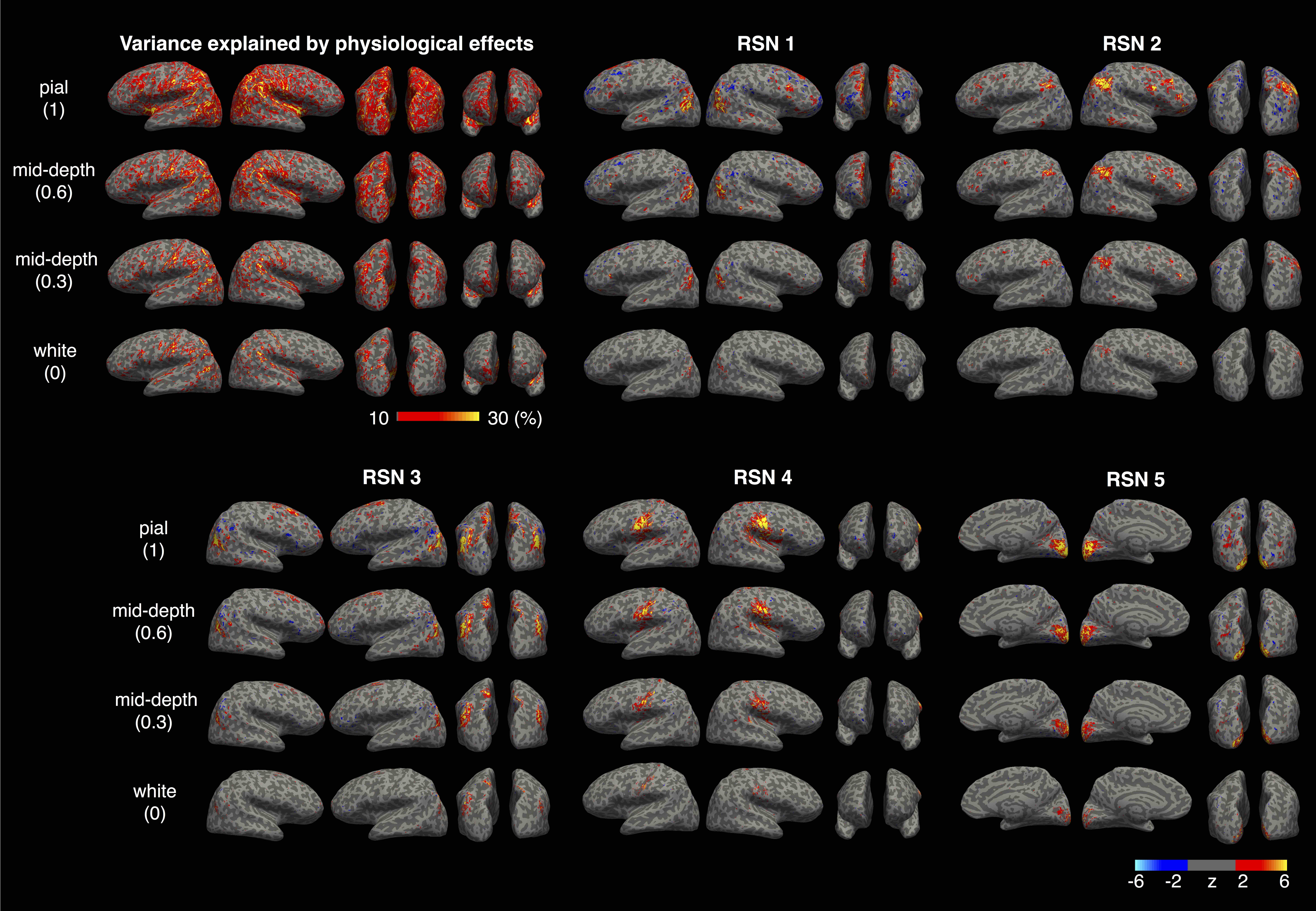
As expected, fractional contributions from physiological changes are most pronounced and spatially extensive near the pial surface, with up to a 3-fold increase across cortical depths (Fig. 4). This confirms our premise that physiological noise contributions are strongest at the pial surface.
Key regions within each RSN transcend all cortical depths, with patterns measured from within the parenchyma being more focal than those measured at the pial surface (Fig. 5). This demonstrates that RSNs can be seen away from the structured physiological noise sources at the pial surface, lending further evidence that these RSNs possess a neuronal origin.
Conclusion
Here, we show that cross-cortical-depth lag patterns show promise for automatically identify neural and non-neural fluctuations. Preliminary characterization of physiological effects and RSFC across cortical depths suggest that excluding voxels intersecting the pial surface can reduce physiological effects and improve neuronal/spatial specificity.Acknowledgements
This work was supported in part by the NIH NIBIB (grants P41-EB015896, and R01-EB019437), by the BRAIN Initiative (NIH NIMH grants R01-MH111438 and R01-MH111419), and by the MGH/HST Athinoula A. Martinos Center for Biomedical Imaging; and was made possible by the resources provided by NIH Shared Instrumentation Grants S10-RR023043 and S10-RR019371.References
[1] Yu X, Glen D, Wang S, Dodd S, Hirano Y, Saad ZS, Reynolds R, Silva AC, Koretsky AP, 2012. Direct imaging of macrovascular and microvascular contributions to BOLD fMRI in layers IV-V of the rat whisker-barrel cortex. Neuroimage. 59(2):1451-1460.
[2] Lewis LD, Setsompop K, Rosen BR, Polimeni JR, 2018. Stimulus-dependent hemodynamic response timing across the human subcortical-cortical visual pathway identified through high spatiotemporal resolution 7T fMRI. Neuroimage. 181:279-291.
[3] Zaretskaya N, Fischl B, Reuter M, Renvall V, Polimeni JR, 2018. Advantages of cortical surface reconstruction using submillimeter 7 T MEMPRAGE. Neuroimage. 165:11-26.
[4] Beckmann CF, DeLuca M, Devlin JT, Smith SM. Investigations into resting-state connectivity using independent component analysis. Philos Trans R Society of London B: Biological Sciences. 2005;360(1457):1001–1013.
[5] Corbin, N., Todd, N., Friston, K.J., Callaghan, M.F., 2018. Accurate modeling of temporal correlations in rapidly sampled fMRI time series. Hum Brain Mapp.
[6] Glover, G. H., Li, T. Q., & Ress, D. (2000). Image‐based method for retrospective correction of physiological motion effects in fMRI: RETROICOR. Magnetic Resonance in Medicine: An Official Journal of the International Society for Magnetic Resonance in Medicine, 44(1), 162-167.
[7] Chang, C., Cunningham, J. P., & Glover, G. H. (2009). Influence of heart rate on the BOLD signal: the cardiac response function. Neuroimage, 44(3), 857-869.
[8] Beissner F, Schumann A, Brunn F, Eisenträger D, Bär K-J, 2014. Advances in functional magnetic resonance imaging of the human brainstem. Neuroimage. 86:91-98.
Figures