0557
Non-negative mean apparent propagators using sum-of-squares optimization: MAP+1University of Copenhagen, Copenhagen, Denmark, 2Linköping University, Linköping, Sweden
Synopsis
Mean apparent propagator (MAP) MRI provides an efficient representation of the three-dimensional q-space signal that is used to characterize the diffusional characteristics of the tissue. The standard implementation of MAP-MRI imposes soft constraints, leaving the possibility for negative values of the propagator; we observed that such physically unjustifiable reconstructions are widespread throughout the brain. Here, we introduce a new framework based on sum-of-squares optimization that guarantees the non-negativity of the reconstructed propagator, thus yielding improved estimates of the propagator as well as the scalar measures derived from it.
Introduction
The mean apparent propagator (MAP)10 MRI technique employs a computationally efficient basis for diffusion-weighted MRI data that enables the reconstruction of the full diffusion propagator, and is being increasingly used to study diffusion characteristics within the brain. A valid set of reconstructed MAP parameters corresponds to a physically plausible, globally non-negative propagator, which all current implementations encourage by enforcing non-negativity constraints in a finite set of points. In this work, we show that such soft constraints are generally insufficient, and result in estimation biases that can exceed variations due to the investigated process (e.g. development, aging) or pathology in group studies. To prevent such biases, we propose a new set of constraints for MAP-MRI that guarantee non-negativity of the propagator globally, based on sum-of-squares constraints that have previously been applied successfully to the cumulant expansion2. The use of these stricter constraints should increase the reliability of MAP-MRI-derived measures in clinical studies.Methods
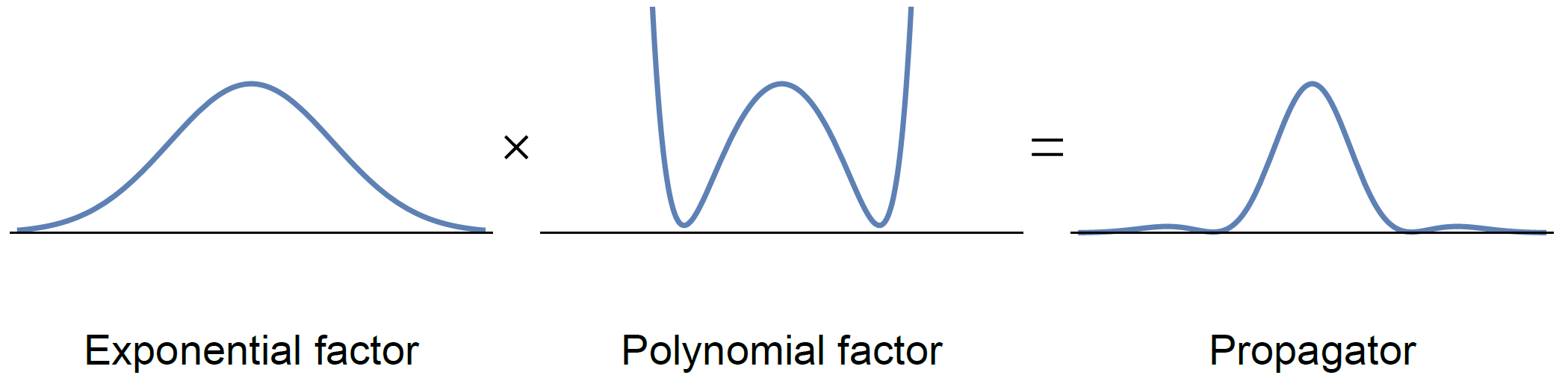
The propagator associated with a MAP fit is given by the product of an exponential factor, typically constructed to correspond to the expected Gaussian behavior described by e.g. DTI, and a polynomial factor that corrects for non-Gaussian effects, cf. Fig. 1. Because the exponential factor is guaranteed to be positive everywhere, non-negativity of the propagator is entirely determined by the polynomial factor. To ensure non-negativity of this polynomial factor—and thus of the entire mean apparent propagator—we propose to use sum-of-squares optimization5.
The central idea behind sum-of-squares optimization is that non-negativity of a polynomial is trivially guaranteed if it can be written as a sum of squared polynomials. Thanks to the denseness results of Lassere4 we know that these sum-of-squares constraints are a practically useful substitute for the general non-negativity constraint, and the works of Parrilo5 and others then show that these constraints can be verified and imposed in practice using semidefinite programming1. In the following experiments we use the semidefinite programming solver SDPA9 (default settings) to validate the sum-of-squares condition on the even order $$$K$$$ polynomial factor
$$\sum_{k=0,2,\ldots,K} \, \sum_{n_1 + n_2 + n_3 = k} \frac{a_{n_1 n_2 n_3}}{\sqrt{2^k \, n_1! n_2! n_3!}} H_{n_1}(r_1) H_{n_2}(r_2) H_{n_3}(r_3),$$
where $$$a_{n_1 n_2 n_3}$$$ are the parameters, $$$n_1$$$, $$$n_2$$$, and $$$n_3$$$ are non-negative integers, and $$$H_n$$$ is the $$$n$$$th order Hermite polynomial. We do this for an entirely unconstrained MAP reconstruction and for the soft constrained (standard) MAP implementation provided by the TORTOISE 3.1.3 package3,6, computed for a subject in the MGH HCP7,8 adult diffusion database. This data set has $$$1.5 \, \mathrm{mm}$$$ isotropic voxels, and $$$b = \{1, 3, 5, 10\} \, \textrm{ms}/\mu\textrm{m}^2$$$ shells with $$$\{64, 64, 128, 256\}$$$ measurements respectively, as well as $$$10$$$ $$$b = 0 \, \textrm{ms}/\mu\textrm{m}^2$$$ images. To distinguish between the soft constrained implementation and our strict sum-of-squares constrained implementation, we will refer to the latter as MAP+. The integral probability constraint10 that is typically included in MAP is obsolete in MAP+ and can be omitted. For the considered orders, the running time for our current implementation of MAP+ is comparable to that of standard MAP.
Results
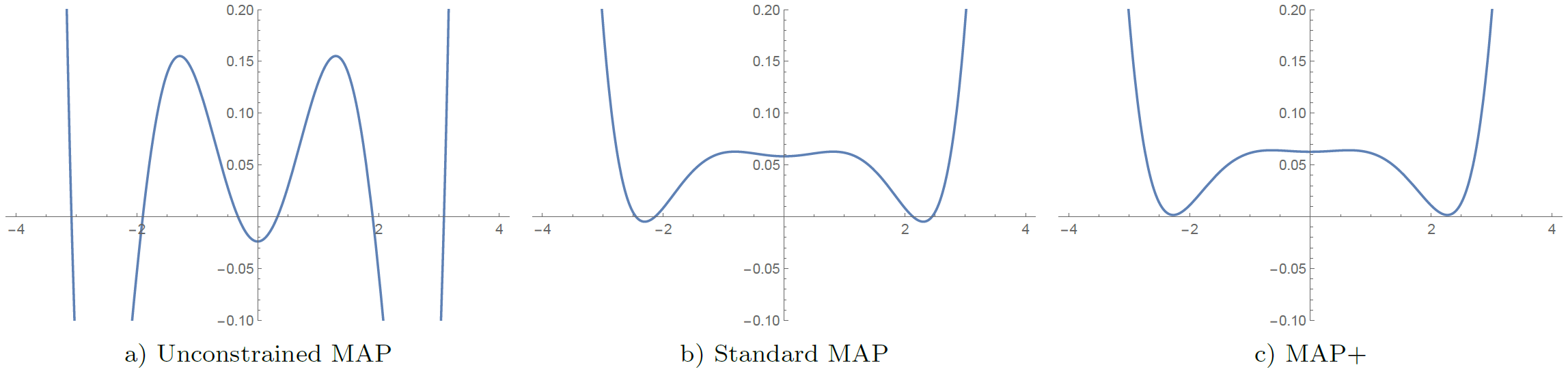
We illustrate the advantage of MAP+ over standard MAP in Fig. 2, where we also compare MAP+ to an entirely unconstrained reconstruction of the propagator. Of the three reconstruction algorithms, only MAP+ produces a polynomial factor / propagator that is non-negative everywhere. Despite their seemingly enormous differences, the difference in the residuals between MAP+ and unconstrained MAP is only a few percent, so the fits are of comparable quality.
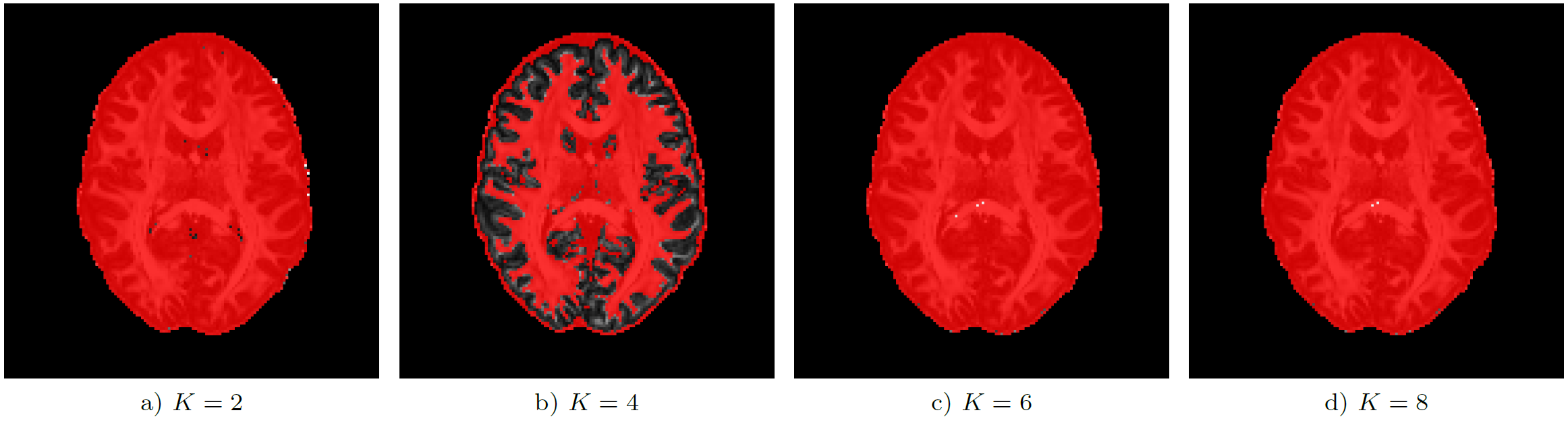
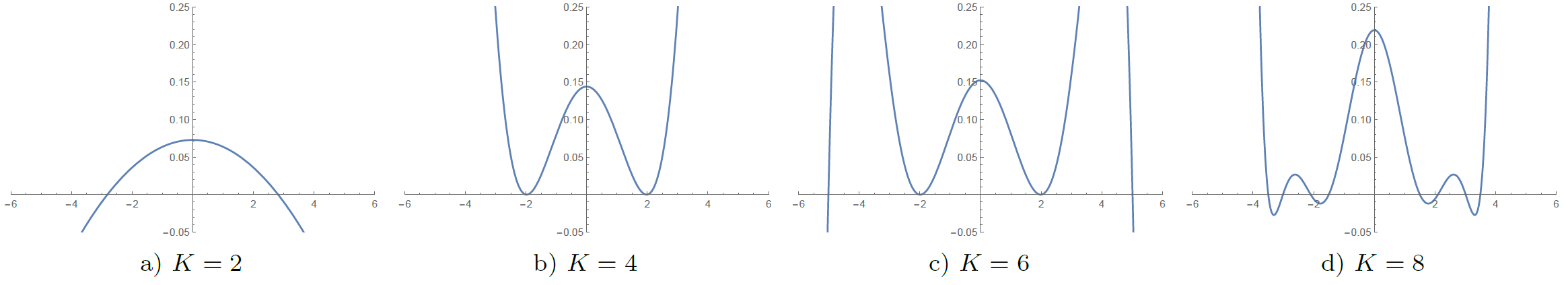
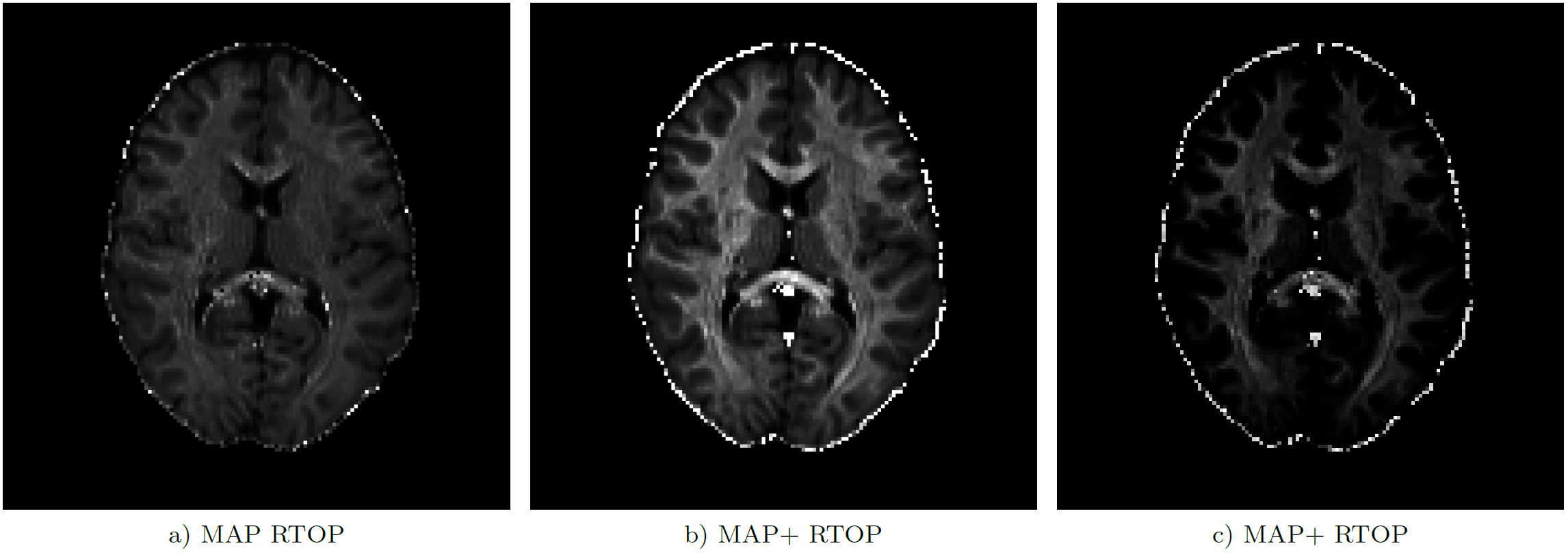
Analysis of the full data set shows that global non-negativity of the propagator is violated in large portions of the image data. Fig. 3 shows the location of these constraint violations in an axial slice, where we identified propagators with negative regions in $$$99.4 \%$$$ ($$$K=2$$$), $$$57.1 \%$$$ ($$$K=4$$$), $$$99.8 \%$$$ ($$$K=6$$$), and $$$99.8 \%$$$ ($$$K=8$$$) of the voxels inside the brain mask ($$$439 248$$$ voxels). The polynomial factor plots in Fig. 4 give some insight into the cause of the observed percentages. We finally note that these violations can have a significant effect on commonly used scalar measures, cf. Fig.5.
Conclusion
Preliminary results on a single data set suggest that the usual constraints for MAP-MRI are quite insufficient—independent of the maximum model order we observed violated constraints in nearly $$$100 \%$$$ of white matter voxels. The resulting bias depends on the specific application but can be significant—the proposed MAP+ approach using sum-of-squares constraints completely nullifies this bias. The computational cost of the MAP+ method is currently being investigated, but is expected to be comparable to the standard MAP approach.Acknowledgements
The authors gratefully acknowledge the Villum Foundation for financial support. Data were provided in part by the McDonnell Center for Systems Neuroscience at Washington University, and by the Human Connectome Project, MGH-USC Consortium (Principal Investigators: Bruce R. Rosen, Arthur W. Toga and Van Wedeen; U01MH093765) funded by the NIH Blueprint Initiative for Neuroscience Research grant; the National Institutes of Health grant P41EB015896; and the Instrumentation Grants S10RR023043, 1S10RR023401, 1S10RR019307.References
[1] S. P. Boyd et al. Convex optimization. 2004.
[2] T. C. J. Dela Haije et al. "The importance of constraints and spherical sampling in diff usion MRI". In: Proceedings of the 26th Annual Meeting of the ISMRM. 2018, p. 5335.
[3] M. O. Irfanoglu et al. "TORTOISE v3: Improvements and new features of the NIH di ffusion MRI processing pipeline". In: Proceedings of the 25th Annual Meeting of the ISMRM. 2017, p. 3540.
[4] J. B. Lasserre. "A sum of squares approximation of nonnegative polynomials". In: SIAM Review 49.4 (2007), pp. 651-669.
[5] P. A. Parrilo. "Semidefi nite programming relaxations for semialgebraic problems". In: Mathematical Programming 96.2 (2003), pp. 293-320.
[6] C. Pierpaoli et al. "TORTOISE: An integrated software package for processing of di ffusion MRI data". en. In: Proceedings of the 18th Annual Meeting of the ISMRM. 2010, p. 1597.
[7] K. Setsompop et al. "Pushing the limits of in vivo di ffusion MRI for the Human Connectome Project". In: NeuroImage80 (2013), pp. 220-233.
[8] D.C. Van Essen et al. "The Human Connectome Project: A data acquisition perspective". In: NeuroImage 62.4 (2012),pp. 2222-2231.
[9] M. Yamashita et al. A high-performance software package for semide finite programs: SDPA 7. Research Report B-460. Tokyo Institute of Technology, 2010.
[10] E. Özarslan et al. "Mean apparent propagator (MAP) MRI: A novel diff usion imaging method for mapping tissue microstructure". In: NeuroImage 78 (2013), pp. 16-32.
[11] E. Özarslan et al. "Simple harmonic oscillator based reconstruction and estimation for one-dimensional q-space magnetic resonance (1D-SHORE)". In: Excursions in Harmonic Analysis, Volume 2. 2013, 373-399.
Figures