0533
Modulation of resting-state functional MRI signal by astrocyteAkira Sumiyoshi1,2, Satoshi Ikemoto1, Elliot A. Stein1, Yihong Yang1, and Hanbing Lu1
1National Institute on Drug Abuse, National Institutes of Health, Baltimore, MD, United States, 2National Institutes for Quantum and Radiological Science and Technology, Chiba, Japan
Synopsis
Although resting-state functional MRI (rs-fMRI) is extensively used to study brain circuitry, recent animal studies suggest a non-neuronal origin of the rs-fMRI signal. We hypothesized that astrocytes may play an important role in rs-fMRI signal. We used chemogenetic technology to selectively activate astrocytes (increasing Ca2+ levels) and recorded rs-fMRI signals in lightly anesthetized rats. Chemogenetic activation of astrocytes following 0.1 mg/kg of clozapine injection induced signal intensity changes and reduced functional connectivity. These in vivo results are consistent with previous brain slice studies, confirming a potentially important role of astrocytes in rs-fMRI signals.
Introduction
Resting-state functional MRI (rs-fMRI) is a neuroimaging method that assesses the strength of functional interactions between brain regions when a subject is not engaged in an explicit task. We have previously identified a robust rs-fMRI circuit between the left and right striatum in lightly anesthetized rats.1 However, simultaneously recorded local field potentials (LFPs) and rs-fMRI signals were only weakly correlated (r < 0.1). A recent optical imaging study in awake mice also reported weak correlations between LFPs and hemodynamic signals during periods of “true” rest in the absence of external stimuli and body movements.2 These results suggest a non-neuronal origin of the rs-fMRI signal. Astrocytes, a type of glial cell, are essential for the supply of energy metabolites to neurons and, by means of their endfoot processes, provide a physical link to the vasculature. We thus hypothesized that astrocytes may directly regulate the vasculature and may play an important role in mediating neurovascular coupling. The aim of this study is to investigate the role of astrocytes in rs-fMRI signal using chemogenetic technology (DREADD; designer receptors exclusively activated by designer drugs). Specifically, we explored the effects of astrocyte-specific DREADD enhancement of calcium activity in striatal astrocytes on the rs-fMRI signal.Methods
In 12 male Sprague-Dawley rats (373–541 g), the active DREADD AAV5-GFAP-hM3D(Gq)-mCherry
was microinjected unilaterally into the striatum while AAV5-GFAP-eGFP was
microinjected as a control into the contralateral striatum (Fig. 1A). The virus
vectors were obtained from Duke Viral Vector Core.3 Left and right sides
were counter-balanced across subjects. After three weeks period for the expression
of DREADD receptors, multiple DREADD-fMRI recordings were conducted from the
same subjects with a one-week interval between data acquisitions (Fig. 1B). Clozapine
(0.1 or 0.5 mg/kg) or clozapine N-oxide (3.0 mg/kg) was used as DREADD agonists.4 Rs-fMRI signals (TR = 1.5 s and spatial resolution =
0.547 x 0.547 x 1 mm3) were acquired using a 9.4-T MRI system with a
20-mm diameter surface coil. Rats were lightly anesthetized with 0.02 mg/kg/hr of
Dexmedetomidine and 0.5% of Isoflurane. All procedures were approved by the
NIDA-IRP Animal Care and Use Committee.Results
The selective expression of DREADD receptors in striatal astrocytes were confirmed using immunohistochemistry (Fig. 1C and D). Region-of-interest (ROI) analyses of rs-fMRI signals, based on individual postmortem histological images (Fig. 2A), demonstrated that astrocyte DREADD activation using 0.1 mg/kg of clozapine induced intensity changes in the rs-fMRI signal that started about 2 min after drug administration and lasted for approximately 20 min (Fig. 2B). In contrast, no changes in signal were seen following 0.5 mg/kg of clozapine or 3.0 mg/kg of clozapine N-oxide injection (Fig. 2C). At the group level there was a significant difference in mean rs-fMRI signal (p < 0.001) between the DREADD and control side ROIs (Fig. 3A). The rs-fMRI signals after DREADD activation showed a bi-phasic response without a few exceptions that showed only an increase or decrease response (Fig. 3B). The rs-fMRI connectivity strength between the DREADD and control side ROIs was significantly reduced after unilateral DREADD activation (one-way ANOVA, p < 0.01) (Fig. 3C).Discussion
Previous brain slice experiments demonstrated that astrocytes are capable of dilating and constricting adjacent arterioles depending on the Ca2+ level of astrocytic endfoot processes and activation of Ca2+ dependent BK channels.5 Brain slice experiments also demonstrate that astrocytes produce a variety of vasodilators and vasoconstrictors (e.g. prostaglandins, epoxyeicosatrienoic acids, and 20-hydroxyeicosatetraenoic acids) via an increase of intracellular Ca2+ levels.6 The rs-fMRI results demonstrated herein using cell-specific DREADD technology are consistent with these previous slice studies, confirming a potentially bidirectional role of astrocytes in generating the rs-fMRI signal.7 Several technical issues need to be considered. First, the rs-fMRI results were not consistent across subjects (Fig. 3B), suggesting differences in cells that were infected and their local circuits, different number of cells, different state of anesthesia, or ongoing brain processes as modulated from other brain regions and circuits. Second, the direct in vivo relationships between astrocytic Ca2+ levels and rs-fMRI signals were not measured, making difficult to infer the temporal dynamics of astrocytic Ca2+ levels compared with rs-fMRI signals. Further technical refinements will be required in the future study.Conclusion
We demonstrated a causal relationship between astrocytic activation and rs-fMRI signals using chemogenetic technology. Upon the selective activation of astrocytes, rs-fMRI signals were fluctuated and the functional connectivity between hemispheres was perturbed. However, several technical issues need to be addressed in the future study to further investigate the role of astrocytes in rs-fMRI signal.Acknowledgements
This work was supported by the Japan Society for the Promotion of Science Overseas Research Fellowships (to AS) and the Intramural Research Program of the National Institute on Drug Abuse, NIH.References
- Jaime S, Gu H, Sadacca BF, et al. Delta rhythm orchestrates the neural activity underlying the resting state BOLD signal via phase-amplitude coupling. Cereb. Cortex, in press.
- Winder AT, Echagarruga C, Zhang Q, et al. Weak correlations between hemodynamic signals and ongoing neural activity during the resting state. Nat. Neurosci. 2017; 20 (12): 1761–1769.
- Bull C, Freitas KCC, Zou S, et al. Rat nucleus accumbens core astrocytes modulate reward and the motivation to self-administer ethanol after abstinence. Neuropsychopharmacology 2014; 39 (12), 2835–2845.
- Gomez JL, Bonaventura J, Lesniak W, et al. Chemogenetics revealed: DREADD occupancy and activation via converted clozapine. Science 2017; 357 (6350), 503–507.
- Girouard H, Bonev AD, Hannah RM, et al. Astrocytic endfoot Ca2+ and BK channels determine both arteriolar dilation and constriction. Proc. Natl. Acad. Sci. U.S.A. 2010; 107 (8), 3811–3816.
- Attwell D, Buchan AM, Charpak S, et al. Glial and neuronal control of brain blood flow. Nature 2010; 468 (7321): 232–243.
- Gordon GRJ, Howarth C, MacVicar BA. Bidirectional control of blood flow by astrocytes: A role for tissue oxygen and other metabolic factors. Adv. Exp. Med. Biol. 2016; 903, 209–219.
Figures
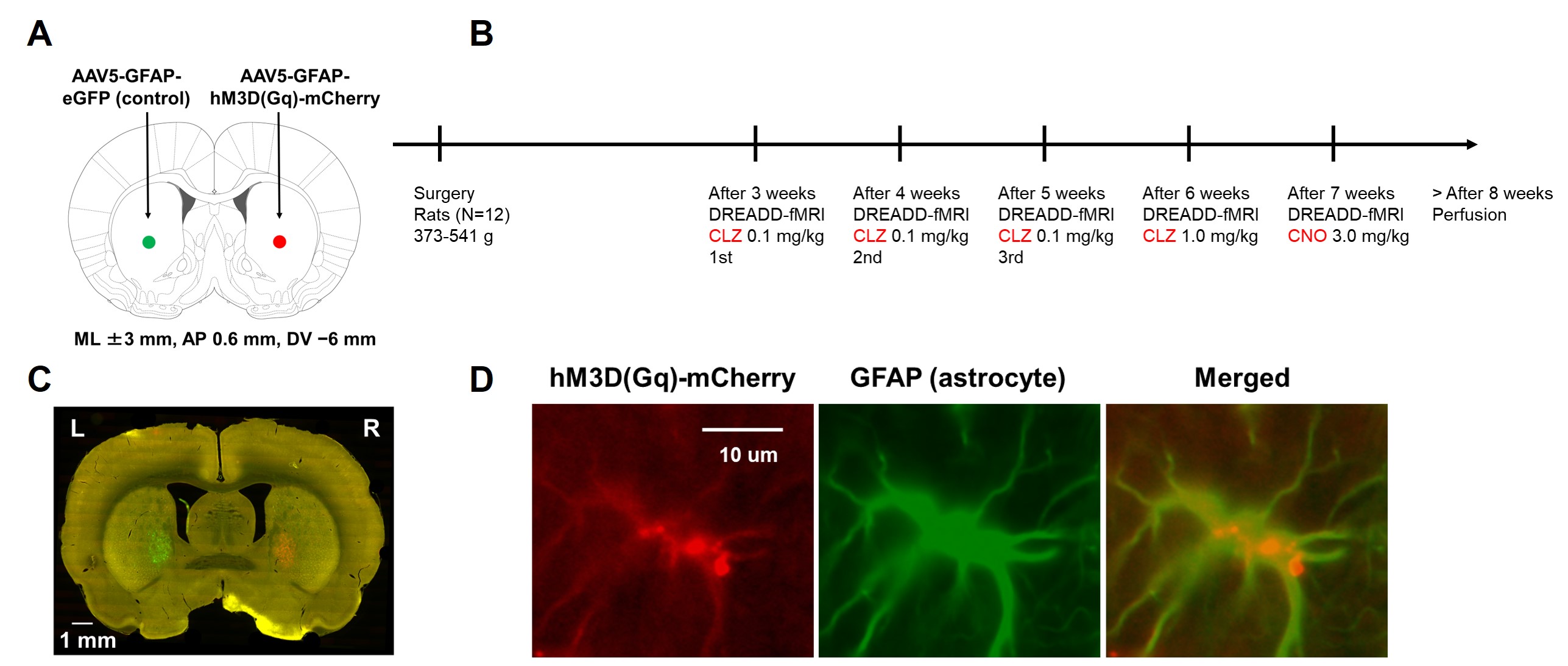
Fig 1. (A) Schematic illustration of microinjection surgery. AAV
vectors were microinjected into left and right striatum. (B) Experimental timeline.
Multiple DREADD-fMRI experiments were conducted from each subject with a
one-week interval. CLZ: clozapine; CNO: clozapine N-oxide. (C) Localization of
DREADD receptor (red) and control eGFP (green). (D) Co-localization of DREADD
receptor (red) with astrocyte marker GFAP (green).
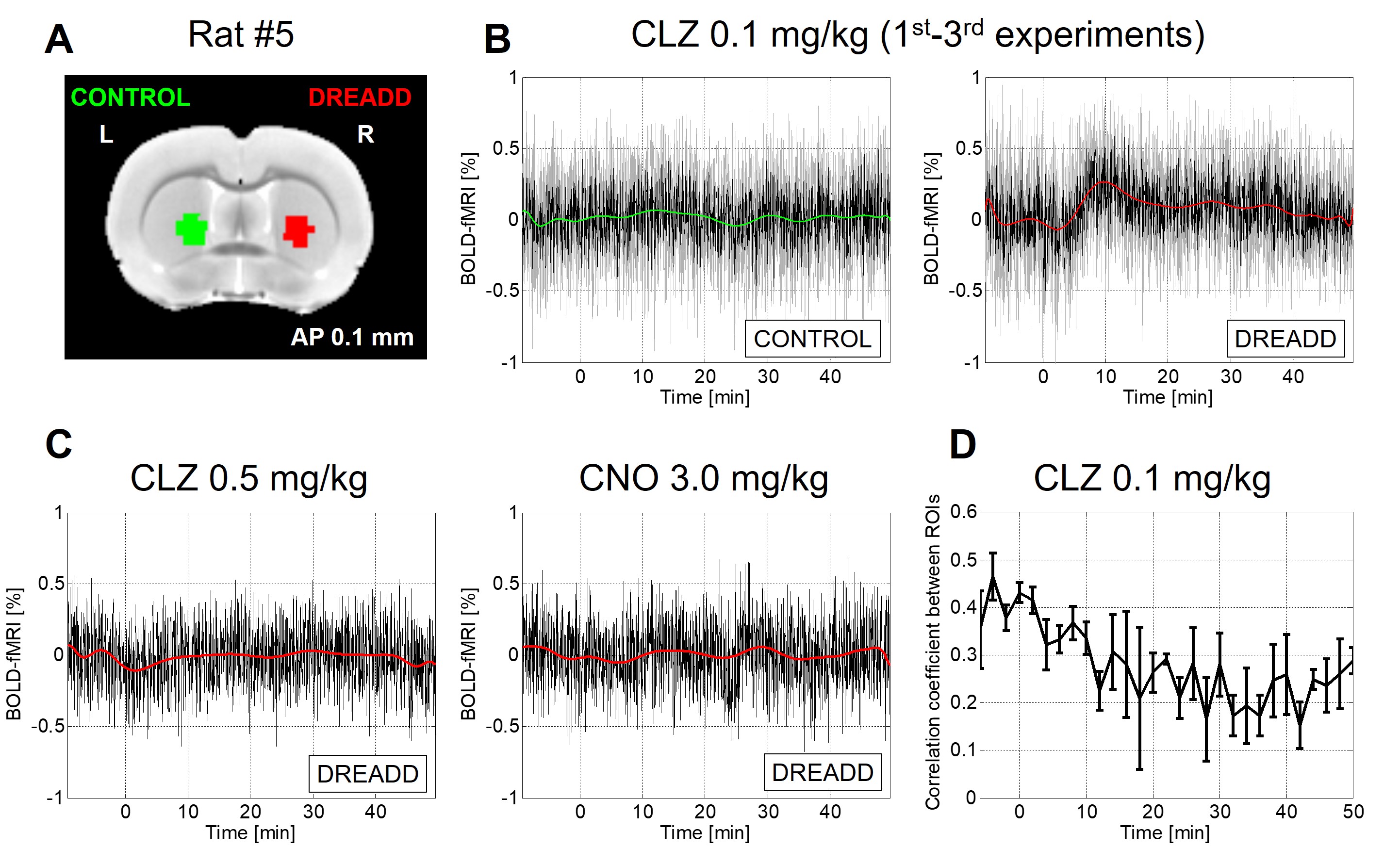
Fig 2. Single-subject
example. (A) ROIs of DREADD and control sides. (B) Comparison of rs-fMRI
signal between DREADD and control side ROIs following 0.1 mg/kg of clozapine
injection. The black line denotes the mean of three experiments and the gray
line denotes each experiment. The green or red line indicates the polynomial
fit curve. (C) Rs-fMRI signal following 0.5 mg/kg of clozapine injection (left) and 3.0 mg/kg
of clozapine N-oxide injection (right). (D) Rs-fMRI connectivity strength between ROIs. Note that correlation coefficient was computed after removing
polynomial fits in DREADD side ROI.
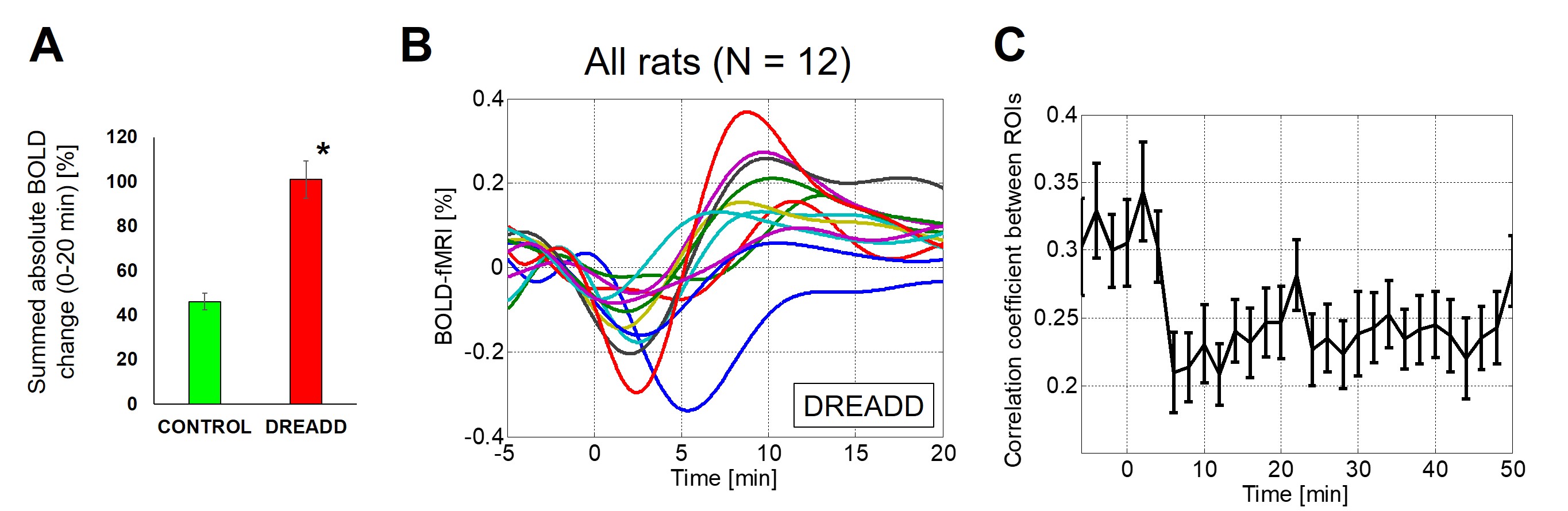
Fig 3. Group data. (A) Summed absolute changes in rs-fMRI signal
between 0 and 20 min following 0.1 mg/kg of clozapine injection were
significantly different between DREADD and control side ROIs (*p < 0.001). (B) Rs-fMRI data of all
subjects following 0.1 mg/kg of clozapine injection (the mean of three
experiments after polynomial fitting). (C) Correlation coefficient between
DREADD and control side ROIs (all subjects, mean +/- sd). Functional
connectivity was perturbed after unilateral DREADD activation (one-way ANOVA, p < 0.01).