0483
Activity of the Pentose Phosphate Pathway Is Increased in Hepatoma Using a Novel Tracer, [2,3-13C2]glucose1Advanced Imaging Research Center, UT Southwestern Medical Center, Dallas, TX, United States, 2Radiology and Internal Medicine, UT Southwestern Medical Center, Dallas, TX, United States
Synopsis
The pentose phosphate pathway (PPP) was investigated in a rat model of hepatoma and results were compared to normal liver and other tissues. A novel and specific tracer of the PPP, [2,3-13C2]glucose, is introduced. The resulting isotopomers are informative because [1,2-13C2]lactate arises only from glycolysis and [2,3-13C2]lactate arises only from the PPP. The PPP was more active in the fed vs. fasted state in most tissues. These results correlated with mRNA expression of key enzymes in the PPP, and flux through both the PPP and glycolysis was substantially increased in hepatoma compared to healthy liver.
Introduction
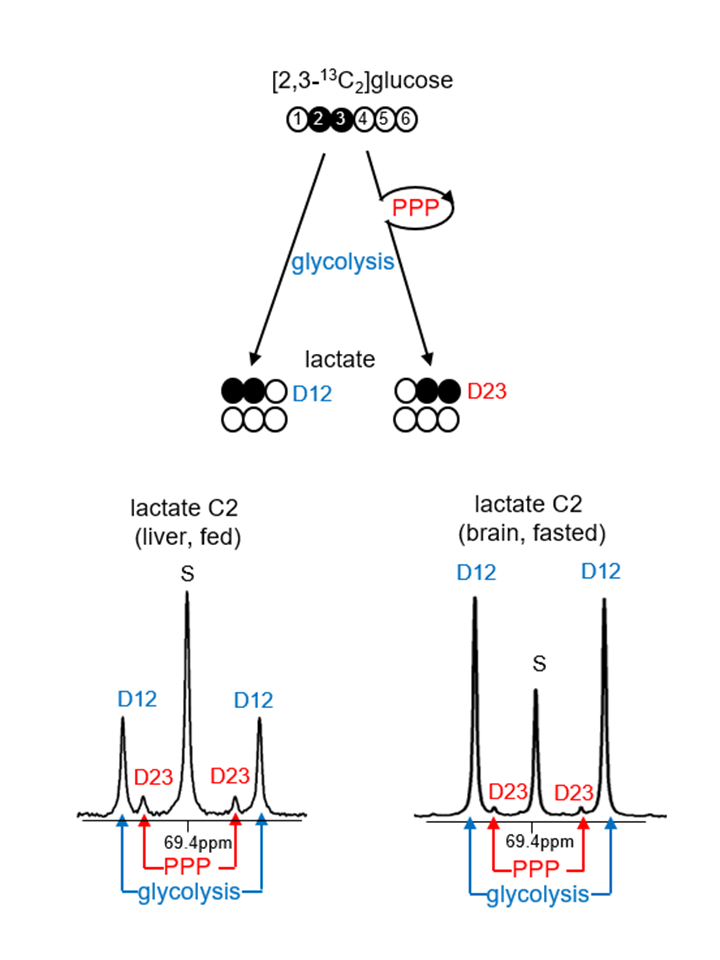
The pentose phosphate pathway (PPP) is essential for biosynthesis of fats in growing cells, antioxidant processes, and production of ribonucleotides for cell reproduction. All of these functions are essential for cancers to grow. Critical enzymes include glucose 6-phosphate dehydrogenase (G6PDH) and transketolase (TKT). Activity of the PPP is thought to be increased in hepatoma (1, 2). Consistent with this hypothesis, aggressive hepatomas are associated with increased glucose uptake by 18FDG PET (3, 4), but PET does not distinguish PPP from glycolysis to lactate. The PPP has been explored using radioactive 14C-labeled tracers (5, 6). Stable isotopes such as [1,2-13C2]glucose have become popular to estimate PPP flux relative to glycolysis (7, 8, 9) because glycolysis yields [2,3-13C2]lactate while the PPP produces [3-13C1]lactate. In principle, this is an attractive method because the two isotopomers are easily quantified, but [3-13C1]lactate via the PPP cannot be distinguished from natural abundance 13C without additional experiments and the assumption that the lactate pool is turning over rapidly. Here we introduce a novel tracer, [2,3-13C2]glucose, to measure the PPP activity in hepatomas. The modification markedly simplifies analysis with the use of NMR. Glycolysis of [2,3-13C2]glucose leads to [1,2-13C2]lactate while the PPP produces [2,3-13C2]lactate. are easily distinguished on 13C NMR (Figure 1), eliminating concerns associated with natural abundance. Furthermore, the critical lactate isotopomer, [2,3-13C2]lactate, cannot arise due to metabolism in the TCA cycle and export of pyruvate, a very active pathway in the liver. The appearance of [2,3-13C2]lactate from [2,3-13C2]glucose is a direct index exclusively of the PPP.Methods
Male Sprague-Dawley rats were studied in three groups: fasted, fed and hepatoma-bearing. The latter group received intrahepatic injections of rat hepatoma cells (N1S1; 1 × 107 cells/100 μL) to establish orthotopic liver tumors which were monitored by MRI to a diameter of ~10 mm. Animals received [2,3-13C2]glucose (99%; 200 mg/kg) intraperitoneally under isoflurane anesthesia. Rats quickly awakened and rested for 60 min before sacrifice under anesthesia. Brain, skeletal muscle, heart and liver of healthy rats (fasted or fed) were harvested, freeze-clamped, and stored for later analysis. From hepatoma-bearing rats, tumors were dissected from liver, and the tumor and surrounding liver tissues were freeze-clamped. Lactate was extracted and spiked with 4,4-dimethyl-4-silapentane-1-sulfonic acid (DSS; 5mM) as a NMR chemical shift and concentration reference; 1H and 13C NMR spectra were acquired at 14.1 T. Real-time quantitative PCR analysis was performed to measure expression of the key PPP enzymes, TKT and G6PDH.Results
PPP activity is enhanced in the heart and liver of fed animals. The PPP flux relative to glycolysis in brain, skeletal muscle, heart and liver of rats was 0-16% based on the ratio of D23/D12 by NMR analysis of lactate C2. This relative PPP flux was higher under a fed compared to a fasted condition in all the organs. Aside from brain tissue, the production of [2,3-13C2]lactate (normalized by DSS; D23/DSS) was also enhanced under a fed condition in skeletal muscle, heart and liver. The enhanced PPP activity was much higher in liver and heart, and the mRNA expressions of G6PDH and TKT were also significantly higher in these two organs under fed conditions.
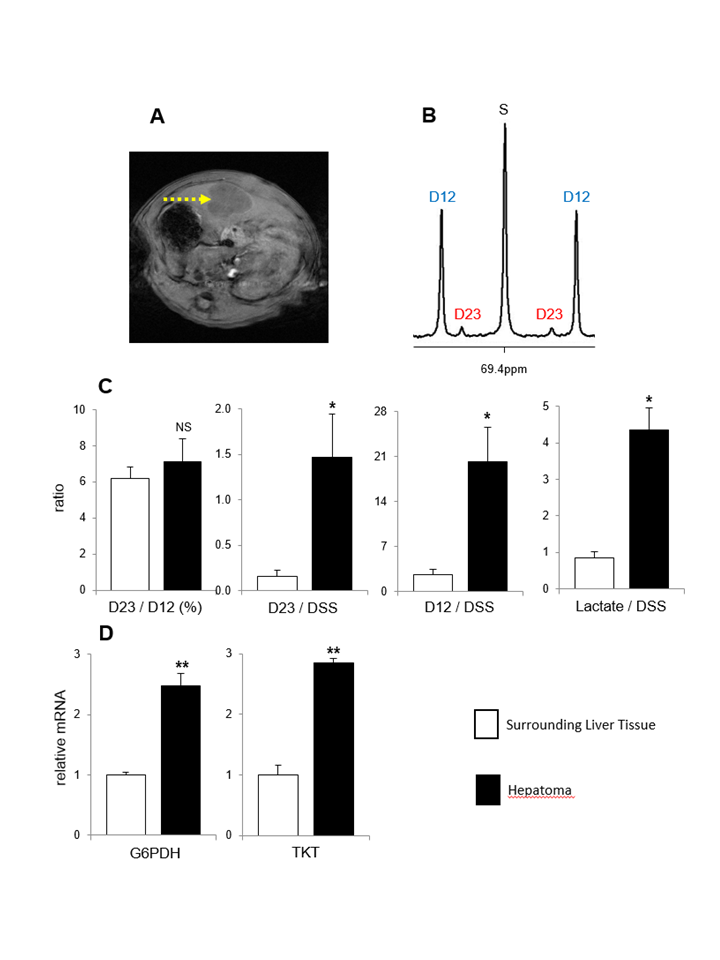
PPP activity and glycolysis are increased in hepatomas. Figure 2 shows MRI of hepatoma and 13C NMR of lactate C2 from hepatoma extracts (Figure 2A-B). The ratio of D23/D12 was not changed, but the levels of both [2,3-13C2]lactate (D23/DSS) and [1,2-13C2]lactate (D12/DSS) were substantially higher in hepatomas compared to surrounding liver tissues; [2,3-13C2]lactate via the PPP was 9-fold and [1,2-13C2]lactate via glycolysis was 8-fold in hepatomas. Also the concentration of lactate measured by 1H NMR was also much higher in hepatomas (Figure 2C). Consistent with the NMR assessment of PPP activity, the mRNA expressions of G6PDH and TKT were higher in hepatomas than the liver surrounding the tumor (Figure 2D).
Discussion and Conclusion
We demonstrated that [2,3-13C2]glucose is a convenient tracer to measure PPP activity. In general, PPP flux was much less active compared to glycolysis regardless of organ or nutritional state, and the PPP measured by NMR correlated with expression of key PPP enzymes, G6PDH and TKT. Both PPP activity and glycolysis were substantially increased in hepatomas, suggesting that the clinical observation of 18FDG uptake reflects both pathways. The use of [2,3-13C2]glucose with monitoring of 13C in lactate is a convenient tool for simultaneously detecting glycolysis and the PPP in cancer.Acknowledgements
P41EB015908References
1. Yin et al. ID1 promotes hepatocellular carcinoma proliferation and confers chemoresistance to oxaliplatin by activating pentose phosphate pathway. J Exp Clin Cancer Res. 2017; 36: 166. 2. Kowalik et al. Emerging Role of the Pentose Phosphate Pathway in Hepatocellular Carcinoma. Front Oncol. 2017; 7: 87. 3. Cho et al. Does 18F-FDG positron emission tomography-computed tomography have a role in initial staging of hepatocellular carcinoma? PLoS One. 2014; 9: e105679. 4. Khan et al. Positron emission tomography scanning in the evaluation of hepatocellular carcinoma. J Hepatol. 2000; 32: 792-7. 5. Scofield et al. The nature of the pentose pathway in liver. J Biol Chem. 1985; 260: 15439-44. PubMed PMID: 3934159. 6. Magnusson I, Chandramouli V, Schumann WC, Kumaran K, Wahren J, Landau BR. Pentose pathway in human liver. Proc Natl Acad Sci U S A. 1988; 85: 4682-5. 7. Lee et al. Mass isotopomer study of the nonoxidative pathways of the pentose cycle with [1,2-13C2]glucose. Am J Physiol. 1998; 274: E843-51. 8. Miccheli et al. Metabolic profiling by 13C-NMR spectroscopy: [1,2-13C2]glucose reveals a heterogeneous metabolism in human leukemia T cells. Biochimie. 2006; 88: 437-48. 9. Marin-Valencia et al. Glucose metabolism via the pentose phosphate pathway, glycolysis and Krebs cycle in an orthotopic mouse model of human brain tumors. NMR Biomed. 2012; 25: 1177-86.Figures