0327
Targeting brain and cognitive aging with multi-modal imaging and connectome topography profiling1Multimodal Imaging and Connectome Laboratory, Montreal Neurological Institute, Montreal, QC, Canada, 2Neuroimaging of Epilepsy Laboratory, Montreal Neurological Institute, Montreal, QC, Canada, 3Brain and Cognition Laboratory, Montreal Neurological Institute, Montreal, QC, Canada
Synopsis
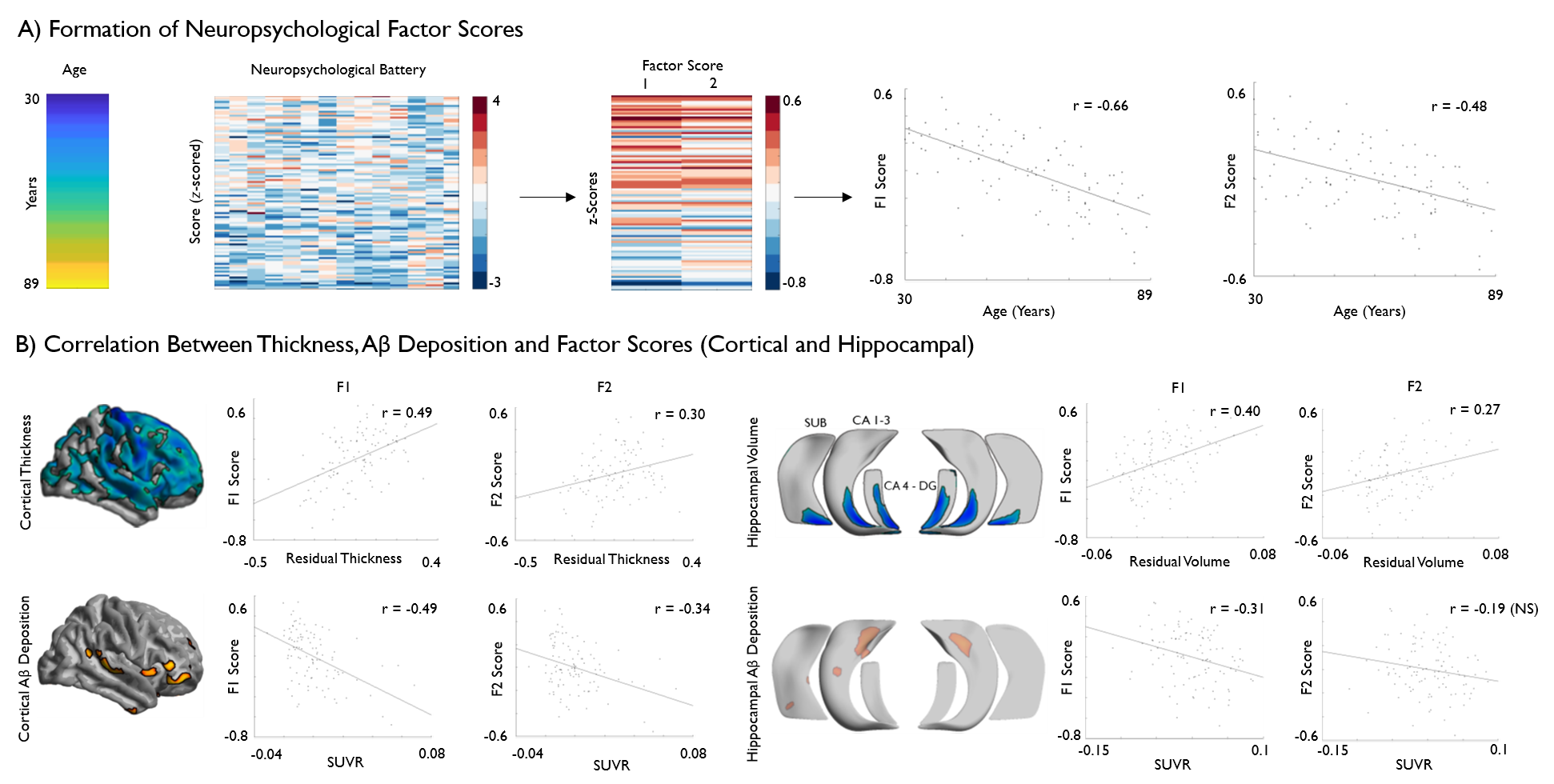
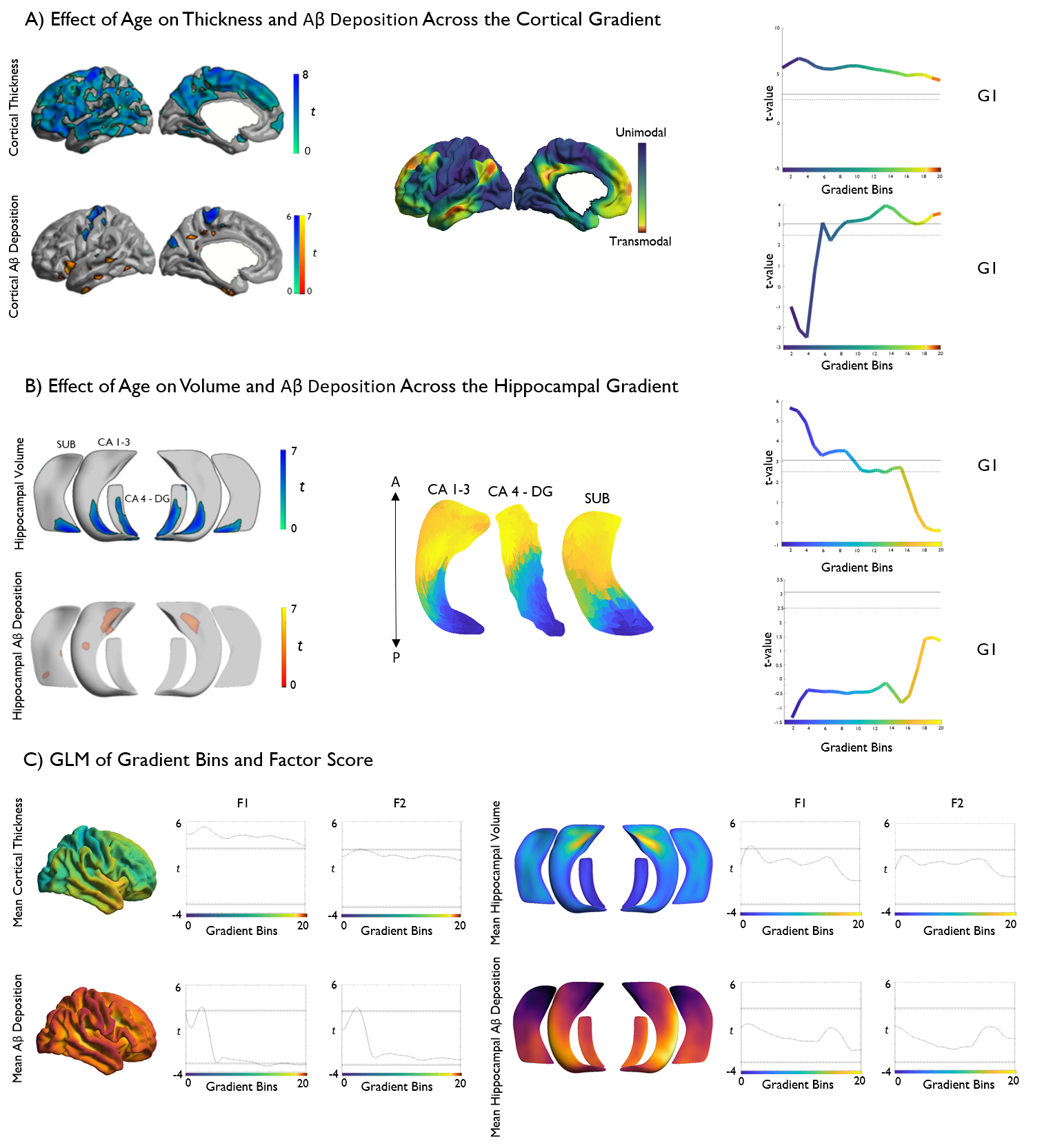
We present an approach to represent and analyze age-related differences in cortical morphology and Aβ uptake based on connectome topography. Studying healthy individuals, we observed age-related reductions in neocortical thickness and atrophy across posterior hippocampal subfields. Additionally, we observed an interplay between aging effects and functional topography in both neocortical and hippocampal regions, with age-related thinning stronger towards unimodal regions and Aβ deposition increasing towards transmodal regions. Similarly, an inverted pattern of volume loss and Aβ deposition was observed along the hippocampal long-axis. Finally, imaging markers were found to predict cognitive performance in a topography-specific manner.
INTRODUCTION
Aging is a complex process involving the accumulation of structural and metabolic changes that ultimately lead to impairments across multiple cognitive domains [1]. With increasing availability of open-access and multi-modal data aggregation and dissemination initiatives, it is now possible to adopt an integrated approach, combining several imaging markers to better understand biological factors contributing to cognitive decline. Here, we present a novel analytical framework to visualize and analyze age-related changes in cortical morphology and amyloid-beta (Aβ) uptake, which represents cortical and hippocampal regions along a compact manifold derived from functional connectome topography, and to relate these data to cognitive profiles in healthy aging. Representing data in this manner assumes a gradual transition between networks, allowing for the effect of age to be accurately represented across the neocortical hierarchy and hippocampal long-axis, thus offering advantages over typical parcellation-based methods.METHODS
Based on a subsample of the open-access Dallas Lifespan Cohort adults (n=102, 69 females, age range=30-89 years), we automatically segmented neocortical and allocortical hippocampal subfield surfaces on high-resolution T1-weighted MRI (3D MPRAGE, TR = 8.1ms, TE = 3.7ms, 1x1x1mm3) obtained in a 3T imaging dataset [2, 3] and co-registered the surface representations to PET-derived Aβ deposition data. Following statistical correction for education and sex, we built linear models to examine effects of age on thickness and partial volume corrected Aβ deposition on neocortical and hippocampal surfaces and to address associations to cognitive profiles. Cognitive profiles pertaining to fluid intelligence and episodic memory were derived from a common factor analysis of the neuropsychological data. In a second step, we reduced data dimensionality using resting-state functional connectome gradient information obtained in an independent dataset [4, 5], which allowed the representation of multimodal imaging profiles relative to neocortical functional topography and hippocampal long axis organization. Both gradients were discretized into 20 bins and the mean thickness and Aβ values were calculated within each bin. We assessed the relationship of age and cognitive factors with neuroimaging markers within each bin with FDR-corrected linear models.RESULTS
Vertex-wise analysis revealed widespread age-related reductions in cortical thickness as well as posterior hippocampal atrophy across all subfields. Age-related increases in Aβ deposition were more confined, occurring primarily in limbic and default mode association cortices. Analysis of age effects along neocortical and hippocampal connectome gradients revealed diffuse thinning across the entire cortical gradient, with stronger effects in unimodal regions, whereas Aβ deposition showed an increase towards transmodal core hubs. A similar inverted effect was observed in the hippocampus, with volume loss observed posteriorly and increased Aβ deposition in anterior subregions. Regarding neuropsychological functioning, regional and gradient-wise findings were found to significantly predict factor-analytical markers of fluid intelligence and episodic memory.DISCUSSION
Our findings build upon existing evidence indicative of structural and metabolic change with advancing age at the neocortical and hippocampal level. Notably, representing data in a novel and compact reference frame centered on functional transitions between systems reveals a striking dichotomy between the structural and metabolic biomarkers along the main axes of neocortical and hippocampal functional organization. These data are consistent with a hypothesized spatial topology of brain aging, wherein posterior structural degradation leads to increased activity in anterior brain regions [6] . Increased neural activity and metabolic demand may in turn lead to increased Aβ deposition, suggesting a structure-metabolic cascade leading to age-related cognitive decline.CONCLUSION
This study presents a novel approach to represent age-related differences in brain structure, metabolism, and cognition. In addition to supporting previous work indicative of structural and metabolic change in the aging brain, the use of a compact analytical framework to relate brain-based biomarkers to macroscale functional systems allowed for novel insights into the interplay between pathological deposits and structural compromise, and how this subsequently impacts upon cognition in the healthy aging population.Acknowledgements
We like to thank the investigators of the DLBS and associated funding sources for making their data available, and INDI/FCP1000 for hosting the imaging data. Reinder Vos de Wael MSc was supported by the McGill Faculty of Medicine. Sara Larivière MSc was supported by a Jeanne Timmins Costello Fellowship. Drs Neda and Andrea Bernasconi were funded by the Canadian Institutes of Health Research (CIHR) and received salary support from the Fonds de la Recherche du Quebec – Santé (FRQS). Dr Boris Bernhardt acknowledges research support from the National Science and Engineering Research Council of Canada (NSERC Discovery-1304413), the Canadian Institutes of Health Research (CIHR FDN-154298), SickKids Foundation (NI17-039), as well as salary support from the Fonds de la Recherche du Quebec - Santé (FRQS Junior 1 Research Scholar).References
[1]. Fjell, AM et al., 2016. Cerebral Cortex, 27(3).
[2]. Fischl, 2012. NeuroImage, 62(2).
[3]. Caldairou et al., 2016. MICCAI.
[4]. Margulies, DS et al., 2016. PNAS, 102(21).
[5]. Vos de Wael, R et al., 2018. PNAS, 115(40).
[6]. Davis et al., 2008. Cerebral Cortex, 18(5).
Figures