0297
Investigating tissue microstructure using NODDI and histology: A chronic injury study on ex vivo macaque spinal cords1Radiology, University of British Columbia, Vancouver, BC, Canada, 2UBC MRI Research Centre, Vancouver, BC, Canada, 3International Collaboration on Repair Discoveries, Vancouver, BC, Canada, 4Department of Spinal Surgery, Nanfang Hospital, Southern Medical University, Guangzhou, China
Synopsis
In this study we sought to understand whether Neurite Orientation Dispersion and Density Imaging (NODDI) could accurately delineate the extent of microstructure damage in an ex-vivo macaque spinal cord. We used histology as a gold standard for delineating the margin between damaged and undamaged tissue. Qualitative analysis exposed disagreements between NODDI maps and histology in areas of damage. This suggests that NODDI may not be appropriate for this model of chronic spinal cord injury.
Introduction
Spinal cord injury (SCI) is a disorder with devastating, often permanent effects on patients. While MRI is commonly used clinically to detect the site of the injury1, the extent of the damage, especially outside of the injury epicentre, remains unclear. NODDI2, a promising diffusion MRI model proposed to provide microstructural information, could be a marker for tissue degeneration. NODDI uses a three compartment model to estimate the intra-cellular (axonal) volume fraction (ICVF), the isotropic volume fraction (ISOVF) and the orientation dispersion (OD) of axons. In this study, we seek to understand whether NODDI can accurately delineate the extent of damage on either side of the injury epicentre in an ex-vivo macaque spinal cord. We use histology as a gold standard for delineating the margin between damaged and undamaged tissue.Methods
MRI Acquisition
8 injured (C5 hemi-contusion, 4 harvested at 24 weeks post injury (wpi), and 4 harvested 48wpi) rhesus macaque spinal cord specimens in 4% paraformaldehyde (hydrolyzed) were scanned using a 35 mm inner-diameter quadrature volume coil in a 7T Bruker Biospec MRI scanner.
A multi-shell 3D diffusion weighted spin echo EPI scan was acquired (TE=37.4ms, TR =250ms, 2 shots, FOV=1.92x1.92 x 4.8cm, matrix=128x128x48, b = 2000, 5000, 7000, and 10000 s/mm2, 81 uniformly distributed diffusion directions per shell3, 4, 5, 4 b=0 s/mm2 scans, total acquisition time = 2hr11min12sec). The diffusion data was preprocessed with MRtrix’s denoising software6, 7. NODDI ICVF and OD maps were then generated using the AMICO software package8.
Histology
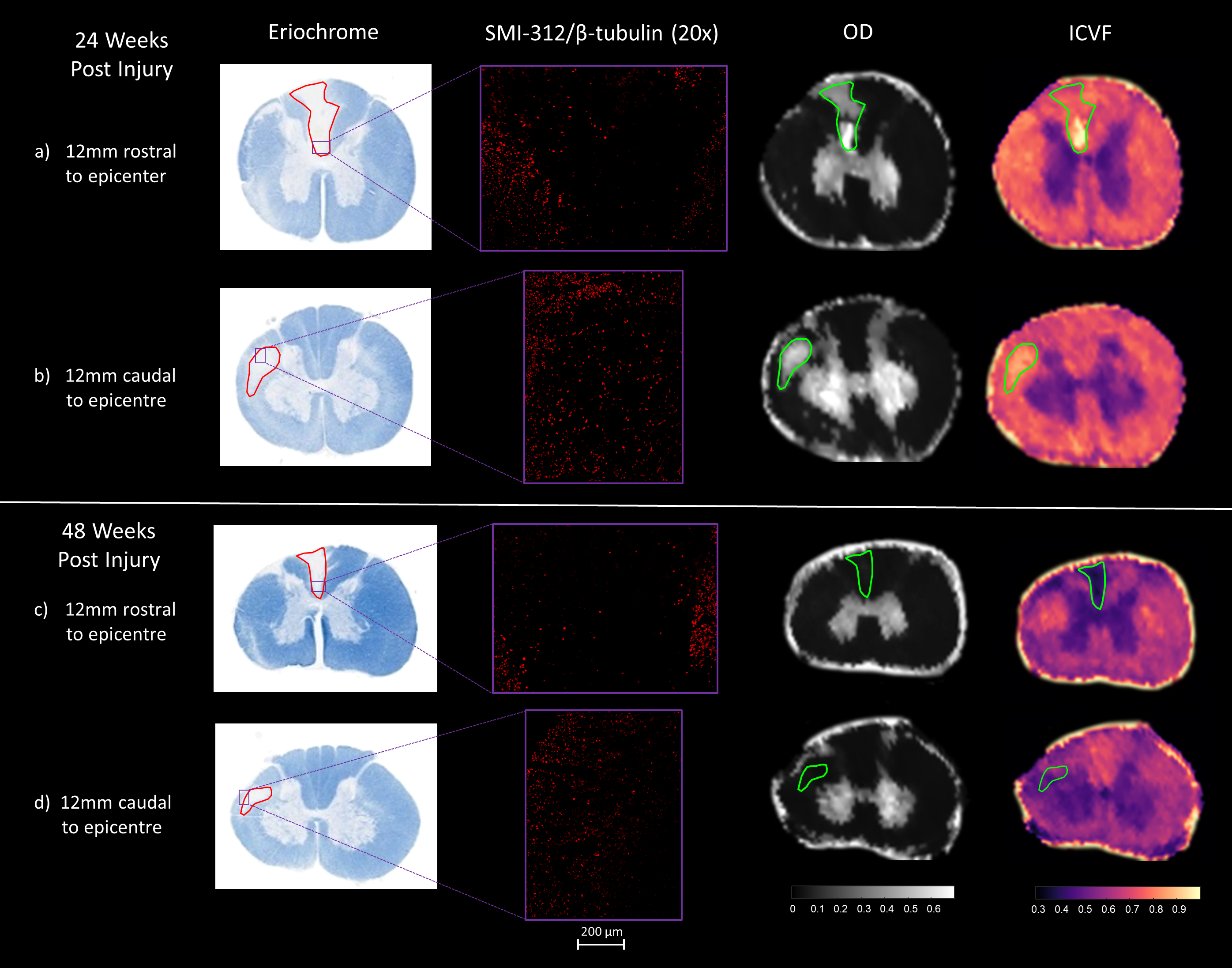
Molds were designed and 3D printed for each specimen to assist registration between MRI data and histology (Figure 1). Tissues slices were immunostained for axon (SMI-312/β-tubulin) and astrocyte (GFAP) content. A separate set of sections were stained for eriochrome (EC). Since EC stains for both intact myelin and myelin debris, an absence of stain highlights areas that are devoid of myelin. ROIs were drawn on these unstained areas to represent an estimate of damaged white matter. NODDI maps were then registered9 to the histology slices with the ROIs overlaid on top. NODDI maps were qualitatively analyzed by comparing the damaged ROIs with the contralateral, uninjured side, of the spinal cord. Additionally, SMI-312/β-tubulin at 20x magnification was used to investigate the axon content in the damaged regions.
Results and Discussion
Figure 2 shows data from two representative animals, one from the 24wpi group, and one from the 48wpi group. The findings described are consistent with the remainder of the animals in each group. For each animal, 2 slice positions, 12 mm rostral and caudal to the injury epicentre, have been studied.
24wpi:
In figure 2a and 2b, OD signal is high in the damaged ROI when compared to white matter on the uninjured contralateral side of the cord. The ICVF signal is also high in the damaged region. This suggests a significant presence of axons; however, few axons can be observed with the SMI-312/β-tubulin axon stain.
48wpi:
In figure 2c and 2d, the ICVF and OD signals are both low within the damaged ROI. This agrees with the SMI-312/β-tubulin axon stain, which indicates a very small number of axons within the damaged region.
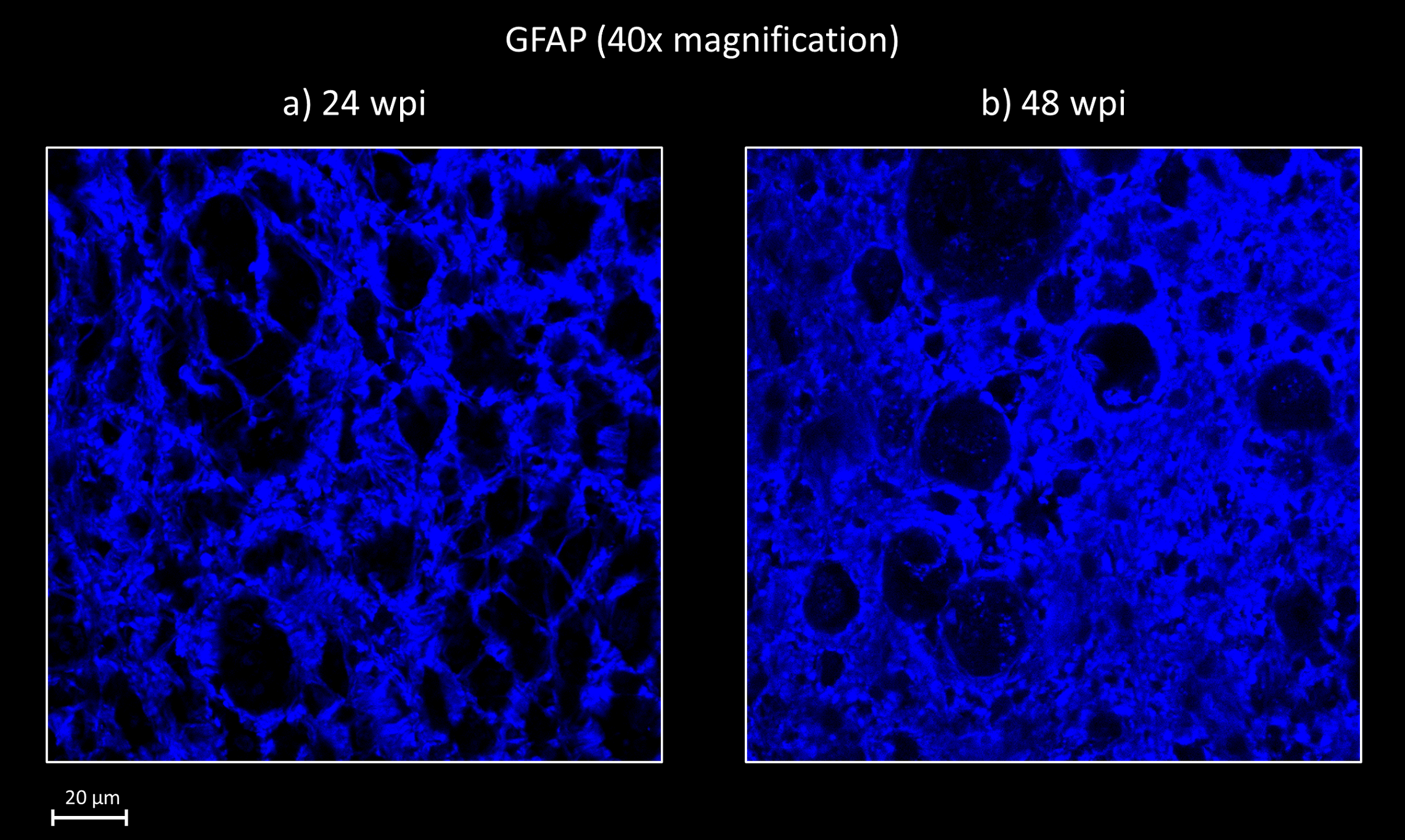
The discrepancy between the 24wpi and 48wpi results suggests that structures classified as the orientationally dispersed axons in the NODDI model at 24wpi, are no longer present at 48wpi. Astrocytes (which normally form a webbed scaffolding around axon bundles) observed with the GFAP stain, led us to hypothesize that at 24wpi (figure 2a), the astrocytes still form a rough boundary around a continuous linear channel that may still produce anisotropic diffusion in the spaces left by the degenerated axons/myelin units. These roughly cylindrical spaces are significantly wider than an axon, which would match our observation of the increased ICVF seen in figure 1a and 1b. In contrast, figure 2b shows that in the 48wpi animal, the processes of the reactive astrocytes are much more densely packed than in the 24wpi animal. We speculate that the astrocytes have filled in much of the spaces once occupied by axons, leaving smaller non-continuous spaces which result in reducing anisotropic diffusion.
Conclusion
With the assistance of histology, we have noticed that the NODDI model of diffusion may misclassify non-axon structures as axons in the chronically injured macaque spinal cord. We suggest that the 3 dimensional shape of astrocytes, in short times after injury, could result in cylindrical spaces with anisotropic diffusion. This could explain the misclassifications, though we acknowledge that future work is required to gain a more robust understanding of the arrangement of astrocytes following axonal degeneration.Acknowledgements
Funding was provided by the Rick Hansen Institute, Vancouver, BC, CanadaReferences
1. Bozzo A, Marcoux J, Radhakrishna M, Pelletier J, Goulet B. The role of magnetic resonance imaging in the management of acute spinal cord injury. J Neurotrauma. 2011;28(8):1401-11.
2. Hui Zhang, Torben Schneider, Claudia A. Wheeler-Kingshott, Daniel C. Alexander. NODDI: Practical in vivo neurite orientation dispersion and density imaging of the human brain. NeuroImage. 2012; 61(4):1000-1016.
3. P. A. Cook, Y. Bai, S. Nedjati-Gilani, K. K. Seunarine, M. G. Hall, G. J. Parker, D. C. Alexander. Camino: Open-Source Diffusion-MRI Reconstruction and Processing. 14th Scientific Meeting of the International Society for Magnetic Resonance in Medicine, Seattle, WA, USA. 2006.
4. Jones DK, Horsfield MA and Simmons A. Optimal strategies for measuring diffusion in anisotropic systems by MRI. Magnetic Resonance in Medicine. 1999; 42(3), 515-525.
5. Jansons KM and Alexander DC. Persistent angular structure: new insights from diffusion magnetic resonance imaging data. Inverse Problems. 2003; 19, 1031-1046.
6. Veraart J, Novikov DS, Christiaens D, Ades-aron B, Sijbers J, Fieremans E. Denoising of diffusion MRI using random matrix theory. NeuroImage. 2016; 142, 394-406.
7. Veraart J, Fieremans E, Novikov DS. Diffusion MRI noise mapping using random matrix theory. Magnetic Resonance in Medicine. 2016; 76(5), 1582-1593.
8. Daducci A, Canales-Rodriguez E, Zhang H, Dyrby T, Alexander D, Thiran JP. Accelerated Microstructure Imaging via Convex Optimization (AMICO) from diffusion MRI data. NeuroImage. 2015; 105, 32-44.
9. Myronenko A, Xubo S. Point-Set Registration: Coherent Point Drift. IEEE Trans. on Pattern Analysis and Machine Intelligence. 2009; 32(12), 2262-2275.
Figures