0223
Is spherical diffusion encoding rotation invariant? An investigation of diffusion time-dependence in the healthy brain1Radiology, Brigham and Women's Hospital, Boston, MA, United States, 2Harvard Medical School, Boston, MA, United States, 3Random Walk Imaging AB, Lund, Sweden, 4Clinical Sciences Lund, Lund University, Lund, Sweden, 55. Danish Research Centre for Magnetic Resonance, Centre for Functional and Diagnostic Imaging and Research, Copenhagen University Hospital Hvidovre, Copenhagen, Denmark, 6Physical Chemistry, Lund University, Lund, Sweden
Synopsis
Recent advances in diffusion weighted MRI have reignited interest in spherical (or isotropic) diffusion encoding. For such encoding to reach high efficiency (minimal echo time), the gradient waveforms have irregular shapes, by design. As such, they lack a well-defined diffusion time and can even be spectrally anisotropic. Most analysis methods based on such encoding assume that diffusion is multi-Gaussian, i.e., that the diffusion is not time-dependent. Since this is a central assumption, we investigate if spherical diffusion encoding is indeed rotation invariant, or if the diffusion time anisotropy has a discernible effect on the diffusion weighted signal in healthy brain.
Introduction
Several recently proposed diffusion MRI techniques employ spherical (or isotropic) diffusion encoding1,2 to probe the microscopic anisotropy and heterogeneity of tissue3-5. To maximize encoding efficiency, the waveforms are often irregular, and therefore lack a straightforward definition of the diffusion time. In 1996, de Swiet and Mitra6 predicted that rotation invariance of spherical encoding could be compromised by time-dependent diffusion7, causing systematic errors in models that ignore time-dependence. Indeed, close inspection of non-conventional encoding waveforms reveals a complex spectral content where diffusion time depends on direction8,9, calling into question the assumption that time-dependence can be ignored. Since ample evidence of time-dependence exists in the brain10,11—albeit at relatively short diffusion times—the reliability of this assumption is imperative to techniques that assume multi-Gaussian diffusion.
In this study, we investigate if spherical diffusion encoding is rotation invariant in a healthy brain by using a waveform that is tuned to exhibit maximal sensitivity to diffusion-time, without impeding its encoding efficiency.
Theory
For axisymmetric diffusion tensors12 with axial $$$(D_\text{A})$$$ and radial eigenvalues $$$(D_\text{R})$$$, the diffusion-weighted signal can be expressed as $$$S=S_0\exp(-bD(\theta))$$$, so that the apparent diffusivity $$$\theta$$$-degrees away from the symmetry axis of the diffusion tensor is$$D(\theta)=\text{MD}+\frac{2}{3}(D_\text{A}-D_\text{R})\cdot{}P_2(\cos\theta)\quad\text{Eq.1}$$where the mean diffusivity $$$\text{MD}=(D_\text{A}+2D_\text{R})/3$$$ and $$$P_2(\cdot)$$$ denotes the second Legendre polynomial. Isotropic diffusion weighting sequences assume a rotationally invariant signal $$$S=S_0\exp(-bD_\text{iso})$$$ where the isotropic diffusivity $$$D_\text{iso}=\text{MD}$$$. However, restrictions may induce time-dependent diffusion and yield a rotation variant inequality between $$$D_\text{iso}$$$ and $$$\text{MD}$$$6,8. The predicted discrepancy can be written as$$\Delta{}D=\frac{2}{3}(D'_\text{A}-D'_\text{R})\cdot{}P_2(\cos\theta)\quad\text{Eq.2}$$where $$$\theta$$$ is now the angle between the low-frequency direction of the spherical encoding and the symmetry axis of the diffusion tensor (Fig.2, right panel), and $$$D'_\text{A}/D'_\text{R}$$$ are the time-dependent apparent axial/radial diffusivities determined by the restriction microstructure and gradient waveform shape. Thus, Eq.2 predicts that $$$\Delta{}D$$$ probed by rotating spherical encoding is proportional to $$$P_2(\cos\theta)$$$ in the presence of anisotropic time-dependence.Methods
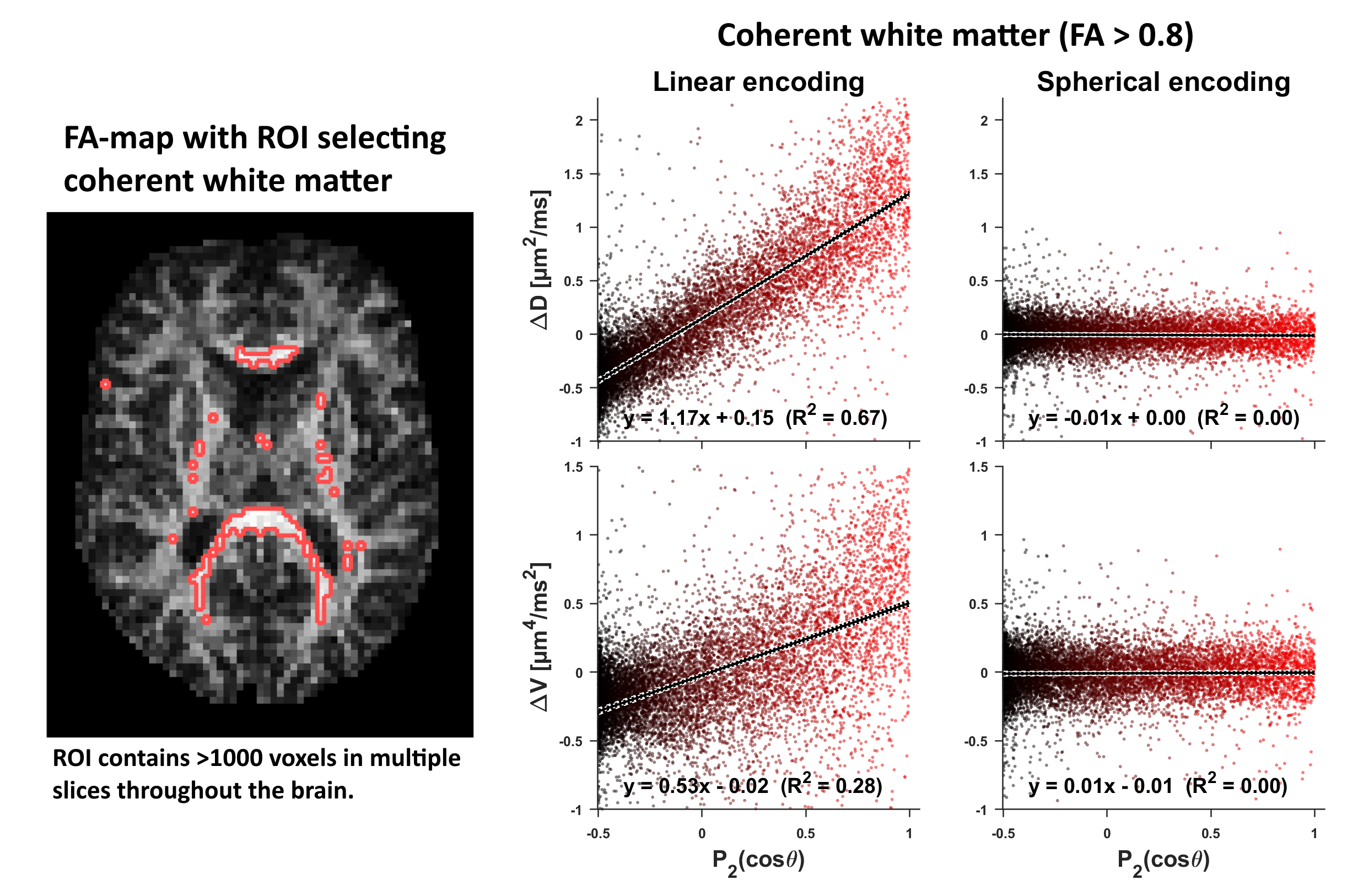
Data was acquired on a 3T (80 mT/m-gradients) in a healthy volunteer, and a liquid crystals phantom with high micro/macro-anisotropy13. Imaging was performed with a prototype spin-echo pulse-sequence where TR/TE=3.2s/91ms, FOV=220x220x60mm3, resolution 2.4mm isotropic, partial-Fourier=7/8, iPAT=2 (Fig.1). Waveforms for spherical encoding were optimized for minimal TE14,15. Importantly, the optimizer was tuned to render maximal diffusion time anisotropy (long diffusion time on x, and short on y and z-axes) without sacrificing the encoding time required to reach the max b-value (Fig.1). Spherical encoding used b=[.1, .7, 1.4, 2] ms/µm2 and ten rotations per shell (rotated so that x-axis pointed along 10 directions). Each series was repeated five times to distinguish variance between and within rotations. Linear encoding used the same b-shells and multiple directions (Fig.1). The number of samples per voxel was 200 for spherical and 82 for linear encoding. Total scan time was 17 min. The fractional anisotropy (FA) and main axis of the diffusion tensor was estimated from conventional diffusion kurtosis analysis16. For each rotation of the spherical encoding, $$$D_\text{iso}$$$ and the isotropic kurtosis $$$V_\text{iso}$$$ (non-normalized5,16) were calculated by fitting the statistical model17$$S_R\approx{}S_0\exp\left(-bD_\text{iso}+\frac{1}{2}b^2V_\text{iso}\right)\quad\text{Eq.3}$$Eq.3 was also used to calculate the mean diffusion and kurtosis (MV) by fitting to signal from all rotations simultaneously. We investigated if the spherical encoding causes rotational variant signal in single voxels of coherent white matter. Furthermore, since single voxel analysis may lack statistical power to detect subtle effects, a comprehensive investigation of rotation dependent diffusivity $$$(\Delta{}D)$$$ and kurtosis $$$(\Delta{}V=V_\text{iso}-MV)$$$ was performed in coherent white matter.Results
Throughout this study, no evidence was found that spherical diffusion encoding depended on rotation. In single voxels of highly coherent white matter (Fig.2), signal was not dependent on rotation; no stratification of $$$S(b)$$$ and the signal variability between and within rotations was on the same order of magnitude. Fig.2 also visualizes the expected signal if rotation variance was present. A comprehensive search in coherent white matter (Fig.3), also revealed no rotation variance of diffusivity or kurtosis. A similar search, restricted to the corticospinal tract and corpus callosum, also indicated that the spherical encoding is invariant to rotation. For verification purposes, the same analysis was performed in simulated and phantom data where time-dependence is known to be null or negligible (Fig.5).Discussion and conclusions
We conclude that the current waveform design indeed produces spherical diffusion encoding with negligible rotation variance, at least in the healthy brain. This greatly simplifies the interpretation of techniques based on the multi-gaussian assumption. However, we recognize that experiments are routinely designed to probe much shorter diffusion times18, and that our findings apply only to waveforms designed to prioritize b-value efficiency, yielding relatively long diffusion times. Furthermore, the current results may not generalize to tissues that differ markedly from the healthy brain, e.g., other organs or tissues affected by disease or development. Investigations of such conditions are forthcoming.Acknowledgements
We thank Siemens Healthcare, Erlangen, Germany, for access to the pulse sequence programming environment. We acknowledge the following research grants NIH P41EB015902, NIH R01MH074794, SSF Framework grant AM13-0090, VR 2016-04482.References
1 Mori, S. & van Zijl, P. Diffusion Weighting by the Trace of the Diffusion Tensor within a Single Scan. Magn. Reson. Med. 33, 41-52, doi:DOI 10.1002/mrm.1910330107 (1995).
2 Wong, E. C. et al. Optimized isotropic diffusion weighting. Magn. Reson. Med. 34, 139-143 (1995).
3 Westin, C. F. et al. Q-space trajectory imaging for multidimensional diffusion MRI of the human brain. Neuroimage 135, 345-362, doi:10.1016/j.neuroimage.2016.02.039 (2016).
4 Topgaard, D. Multidimensional diffusion MRI. J. Magn. Reson. 275, 98-113, doi:10.1016/j.jmr.2016.12.007 (2017).
5 Szczepankiewicz, F. et al. The link between diffusion MRI and tumor heterogeneity: Mapping cell eccentricity and density by diffusional variance decomposition (DIVIDE). Neuroimage 142, 522-532, doi:10.1016/j.neuroimage.2016.07.038 (2016).
6 de Swiet, T. M. & Mitra, P. P. Possible Systematic Errors in Single-Shot Measurements of the Trace of the Diffusion Tensor. J. Magn. Reson. B 111, 15-22 (1996).
7 Stepisnik, J. Time-Dependent Self-Diffusion by Nmr Spin-Echo. Physica B 183, 343-350, doi:Doi 10.1016/0921-4526(93)90124-O (1993).
8 Lundell, H. et al. in Proc. Intl. Soc. Mag. Reson. Med. 25 (Honolulu, Hawaii, 2017).
9 Lundell, H. et al. in Proc. Intl. Soc. Mag. Reson. Med. 26. (Paris, France, 2018)
10 Assaf, Y. et al. AxCaliber: a method for measuring axon diameter distribution from diffusion MRI. Magn. Reson. Med. 59, 1347-1354, doi:10.1002/mrm.21577 (2008).
11 Stanisz, G. J. et al. An analytical model of restricted diffusion in bovine optic nerve. Magn. Reson. Med. 37, 103-111 (1997).
12 Basser, P. J. et al. MR diffusion tensor spectroscopy and imaging. Biophys. J. 66, 259-267, doi:10.1016/S0006-3495(94)80775-1 (1994).
13 Nilsson, M. et al. Liquid crystal phantom for validation of microscopic diffusion anisotropy measurements on clinical MRI 3 systems. Magn. Reson. Med., Submitted, Under minor revision (2017).
14 Sjölund, J. et al. Constrained optimization of gradient waveforms for generalized diffusion encoding. J. Magn. Reson. 261, 157-168, doi:10.1016/j.jmr.2015.10.012 (2015).
15 Szczepankiewicz, F. & Nilsson, M. in Proc. Intl. Soc. Mag. Reson. Med. 26.
16 Jensen, J. H. et al. Diffusional kurtosis imaging: the quantification of non-gaussian water diffusion by means of magnetic resonance imaging. Magn. Reson. Med. 53, 1432-1440, doi:10.1002/mrm.20508 (2005).
17 Yablonskiy, D. A. et al. Statistical model for diffusion attenuated MR signal. Magn. Reson. Med. 50, 664-669, doi:10.1002/mrm.10578 (2003).
18 Does, M. D. et al. Oscillating gradient measurements of water diffusion in normal and globally ischemic rat brain. Magn. Reson. Med. 49, 206-215, doi:10.1002/mrm.10385 (2003).
Figures