0204
Multimodal awake mouse imaging: from two-photon microscopy to BOLD-fMRI1Physique, génie physique et optique, Université Laval, Québec, QC, Canada, 2University of California San Diego, La Jolla, CA, United States, 3Boston University, Boston, MA, United States, 4Martinos Center for Biomedical Imaging, MGH, Harvard Medical School, Charlestown, MA, United States, 5University of Texas at Austin, Austin, TX, United States
Synopsis
Functional Magnetic Resonance Imaging (fMRI) in awake behaving mice is well positioned to bridge the detailed cellular-level view of brain activity, which has become available due to recent advances in microscopic optical imaging and genetics, to the macroscopic scale of human noninvasive observables. Here, we demonstrate Blood Oxygen Level Dependent (BOLD) fMRI in awake mice implanted with chronic transparent cranial ''windows'', compatible with two-photon microscopy, optical imaging, and optogenetic light stimulation. We thus provide a proof of feasibility for multimodal imaging approaches in awake mice, which in the future can be extended to behavioral studies and biomedical applications.
Introduction
Blood Oxygen Level Dependent (BOLD) fMRI is widely used to investigate the human brain, but its interpretation in terms of the underlying microscopic physiology, such as electrical activity of single neurons and hemodynamic activity of single blood vessels, is still under investigation.1 Noninvasive imaging in animal models can play a critical role in physiological underpinning and data-driven modeling of human noninvasive signals, in particular when both micro- and macroscopic measurements can be achieved in the same subject under analogous experimental conditions.
A majority of animal studies are typically performed in anesthetized animals. However, anesthesia can differentially affect neuronal cell types, blood flow and oxygen metabolism, altering neuro-vascular-metabolic coupling and the hemodynamic response.
Here, we provide a protocol and a proof of feasibility for BOLD fMRI in awake mice implanted with chronic glass “cranial windows” that do not significantly deteriorate the quality of fMRI. These windows provide optical access for micro- and “mesoscopic” optical imaging modalities and neuronal stimulation with light (optogenetics 2). Therefore, alternating between the imaging modalities for each subject over the course of weeks to months is possible.
Methods
Mice (N=16) were surgically implanted with a head bar and a glass window sealing a 3-mm round cranial exposure over the barrel cortex (Fig. 1A-B) and left to heal for >21 days. Following behavioral training, they were imaged head-fixed using two-photon microscopy (Fig. 1C-D), spectral optical imaging of blood oxygenation, and BOLD-fMRI, using a custom-made mouse holder (Fig. 2B). In each modality, we measured the hemodynamic response to both sensory stimuli (air puffs to the whiskers at 3-5 Hz for 2s (''event-realted'') and 20s (''blocked'')) as well as optogenetic stimuli (single 100 ms blue light pulse (''event-related''), or repeated at 1Hz for 20s (''blocked''). Optogenetic stimulation allowed for selective stimulation of pyramidal excitatory neurons (in the Emx1-Cre/Ai32 mouse line) or inhibitory interneurons (in VGAT-ChR2(H134R)-EYFP mice)). The workflow for fMRI data processing is shown in Fig. 2A.Results
We first demonstrate that the presence of the window results in minimal artifacts in MRI images (Fig. 1E) while allowing optogenetic stimulation by simply positioning an optical fiber next to the window. In agreement with previous studies that utilized this type of cranial implants for two-photon imaging 3, the window remained clear and transparent for the duration of weeks and months (Fig. 1C) allowing imaging throughout the cortical depth and down to the white matter (Fig. 1D).
On the microscopic scale, two-photon imaging can be used to monitor the diameter of blood vessels which underlie the hemodynamic response. Figure 3A-C illustrates an example of time-resolved imaging of single-vessel dilation, which is a key parameter in detailed models of fMRI signals 4, 5.
Figure 3D show the corresponding mesoscopic changes in blood oxygenation in the same subject under the same stimulus conditions using single-photon CCD-based imaging of hemoglobin-based optical intrinsic signals (OIS) imaging 6-8 to obtain evoked changes in oxyhemoglobin (HbO) and deoxyhemoglobin (HbR) concentrations. These optical measurements provide an easy implementation, low cost proxy for the BOLD-fMRI signal (which results from HbR concentration changes).
Figure 4 demonstrates robust, statistically significant single-subject BOLD response to optogenetic (A-E) and sensory (F-J) stimuli from an awake behaving mouse. Some studies used sedative drugs to mitigate motion artifacts when imaging awake animals 9. In our hands, however, sedation with 1-2 mg/kg chlorprothixene (Sigma-Aldrich) notably slowed down the hemodynamic response kinetics, possibly abolished the undershoot, and evoked a more spatially widespread BOLD-fMRI response (Fig. 5).
Discussion
The results provide a proof-of-principle demonstration for using chronic optical cranial windows in animal BOLD fMRI studies, supporting multimodal and multiscale imaging within one animal. Compared to our published data from rats anesthetized with alpha-chloralose 10, the BOLD signal induced by sensory stimulation in awake mice was ~3 times smaller when normalized by the corresponding averaged arteriolar dilation, highlighting the importance of imaging awake animals in order to exclude any physiological effects of anesthesia when addressing biological questions. Even sedation is not recommended in quantitative hemodynamic and neurovascular coupling studies due to its effect on response dynamics.Conclusion
In the future, sampling multiple physiological parameters in an awake behaving mouse across scales and measurement modalities, including BOLD fMRI, will be instrumental for bridging BOLD fMRI signals induced by a complex behavior to the underlying activity of neuronal circuits. The ability to perform longitudinal studies, while alternating between imaging modalities and manipulating neuronal activity with optogenetic tools, can also facilitate neurophysiological underpinning of spontaneous (“resting state”) hemodynamic fluctuations 11-14 as well as cortical MRI signals in response to clinically-relevant perturbations of brain activity 15.Acknowledgements
We gratefully acknowledge support from the NIH (MH111359, NS057198, U01NS094232, and S10RR029050). M. Desjardins was supported by a postdoctoral scholarship from the Natural Sciences and Engineering Research Council of Canada. K. Kılıç was supported by a postdoctoral fellowship from the International Headache Society and TUBITAK. M. Thunemann was supported by a postdoctoral fellowship from the German Research Council (DFG TH 2031/1).References
1. Uhlirova H, Kilic K, Tian P, Sakadzic S, Gagnon L, Thunemann M, et al. The roadmap for estimation of cell-type-specific neuronal activity from non-invasive measurements. Philos Trans R Soc Lond B Biol Sci. 2016;371(1705).
2. Deisseroth K. Optogenetics: 10 years of microbial opsins in neuroscience. Nat Neurosci. 2015;18(9):1213-25.
3. Goldey GJ, Roumis DK, Glickfeld LL, Kerlin AM, Reid RC, Bonin V, et al. Removable cranial windows for long-term imaging in awake mice. Nat Protoc. 2014;9(11):2515-38.
4. Gagnon L, Sakadzic S, Lesage F, Musacchia JJ, Lefebvre J, Fang Q, et al. Quantifying the microvascular origin of BOLD-fMRI from first principles with two-photon microscopy and an oxygen-sensitive nanoprobe. J Neurosci. 2015;35(8):3663-75.
5. Gagnon L, Sakadzic S, Lesage F, Pouliot P, Dale AM, Devor A, et al. Validation and optimization of hypercapnic-calibrated fMRI from oxygen-sensitive two-photon microscopy. Philos Trans R Soc Lond B Biol Sci. 2016;371(1705).
6. Devor A, Sakadzic S, Yaseen MA, Roussakis E, Tian P, Slovin H, et al. Functional imaging of cerebral oxygenation with intrinsic optical contrast and phosphorescent probes. In: Weber B, Helmchen F, editors. Optical imaging of cortical circuit dynamics. New York: Springer; 2013.
7. Dunn AK, Devor A, Bolay H, Andermann ML, Moskowitz MA, Dale AM, et al. Simultaneous imaging of total cerebral hemoglobin concentration, oxygenation, and blood flow during functional activation. Opt Lett. 2003;28(1):28-30.
8. Dunn AK, Devor A, Dale AM, Boas DA. Spatial extent of oxygen metabolism and hemodynamic changes during functional activation of the rat somatosensory cortex. Neuroimage. 2005;27(2):279-90.
9. Bonder DE, McCarthy KD. Astrocytic Gq-GPCR-linked IP3R-dependent Ca2+ signaling does not mediate neurovascular coupling in mouse visual cortex in vivo. J Neurosci. 2014;34(39):13139-50.
10. Tian P, Teng IC, May LD, Kurz R, Lu K, Scadeng M, et al. Cortical depth-specific microvascular dilation underlies laminar differences in blood oxygenation level-dependent functional MRI signal. Proc Natl Acad Sci U S A. 2010;107(34):15246-51.
11. Murphy MC, Chan KC, Kim SG, Vazquez AL. Macroscale variation in resting-state neuronal activity and connectivity assessed by simultaneous calcium imaging, hemodynamic imaging and electrophysiology. Neuroimage. 2018;169:352-62.
12. Schwalm M, Schmid F, Wachsmuth L, Backhaus H, Kronfeld A, Aedo Jury F, et al. Cortex-wide BOLD fMRI activity reflects locally-recorded slow oscillation-associated calcium waves. Elife. 2017;6.
13. Mateo C, Knutsen PM, Tsai PS, Shih AY, Kleinfeld D. Entrainment of Arteriole Vasomotor Fluctuations by Neural Activity Is a Basis of Blood-Oxygenation-Level-Dependent "Resting-State" Connectivity. Neuron. 2017;96(4):936-48 e3.
14. Ma Y, Shaik MA, Kozberg MG, Kim SH, Portes JP, Timerman D, et al. Resting-state hemodynamics are spatiotemporally coupled to synchronized and symmetric neural activity in excitatory neurons. Proc Natl Acad Sci U S A. 2016;113(52):E8463-E71.
15. Albaugh DL, Salzwedel A, Van Den Berge N, Gao W, Stuber GD, Shih YY. Functional Magnetic Resonance Imaging of Electrical and Optogenetic Deep Brain Stimulation at the Rat Nucleus Accumbens. Sci Rep. 2016;6:31613.
16. Holland D, Kuperman JM, Dale AM. Efficient correction of inhomogeneous static magnetic field-induced distortion in Echo Planar Imaging. Neuroimage. 2010;50(1):175-83.
Figures
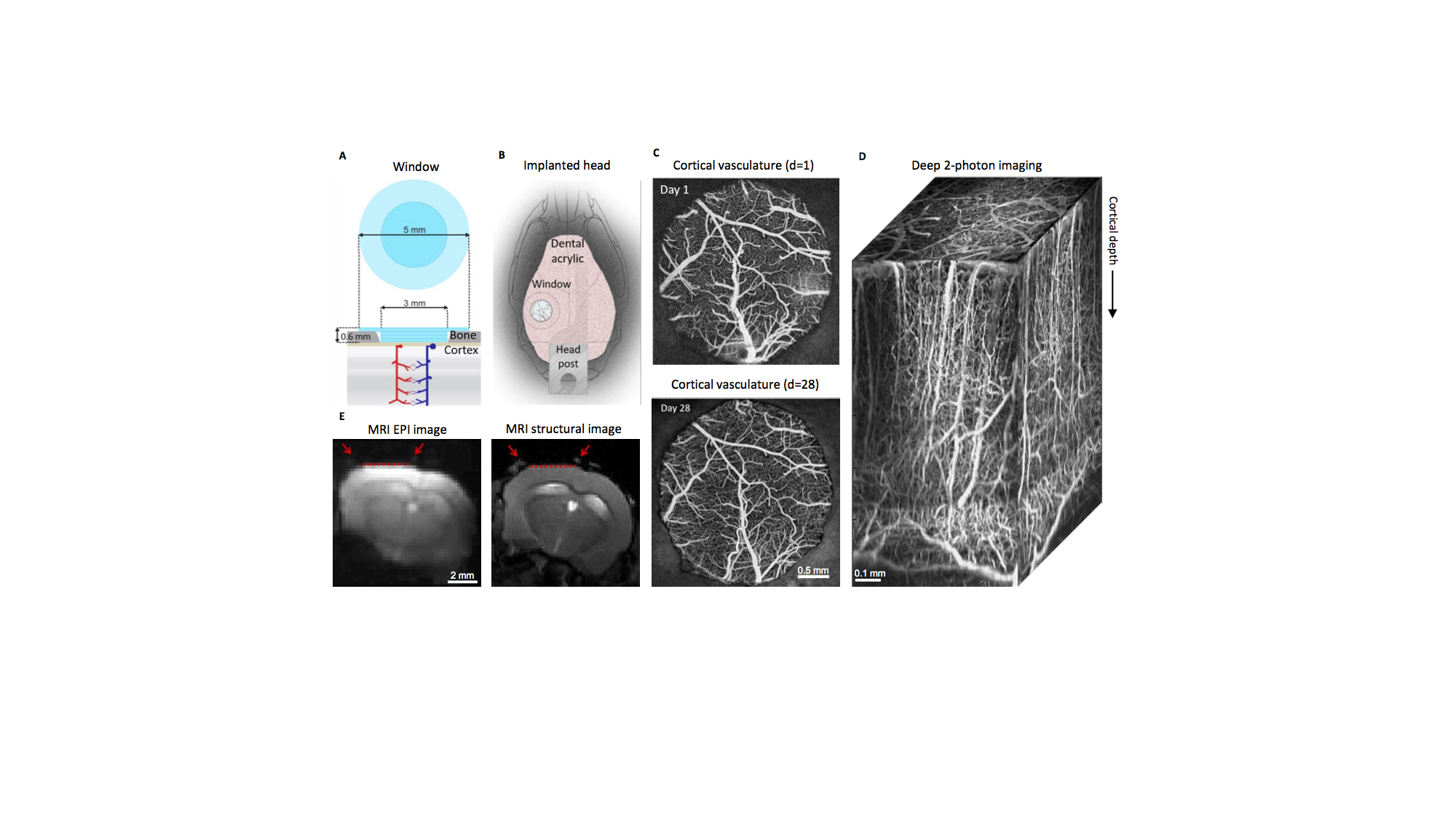
MRI-compatible headpost assembly and image quality across modalities.
A) Borosilicate glass window implant. B) Positioning of window over the barrel cortex and headpost fixed to the contralateral skull. C) Images of brain vasculature through the window from two-photon imaging of fluorescein isothiocyanate labeled dextran injected intravenously, showing preserved integrity of vasculature between days 1 and 28 following surgical implantation. D) Two-photon image stack obtained with intravascular Alexa 680 labeled dextran, illustrating capability of deep imaging. E) Gradient-echo EPI image (left) and structural image (TurboRARE, right). Red arrows point to the glass/bone boundary; red line to the glass/brain boundary.

fMRI methods.
A) Schematics of the protocol for MRI data acquisition and processing. Image distortion was corrected using our previously published method that involves acquisition of spin-echo (SE) echo-planar imaging (EPI) scans with opposite phase encoding polarities 16. B) Custom-made MRI-compatible mouse cradle including functional components for awake mouse imaging with sensory and optogenetic stimulation.
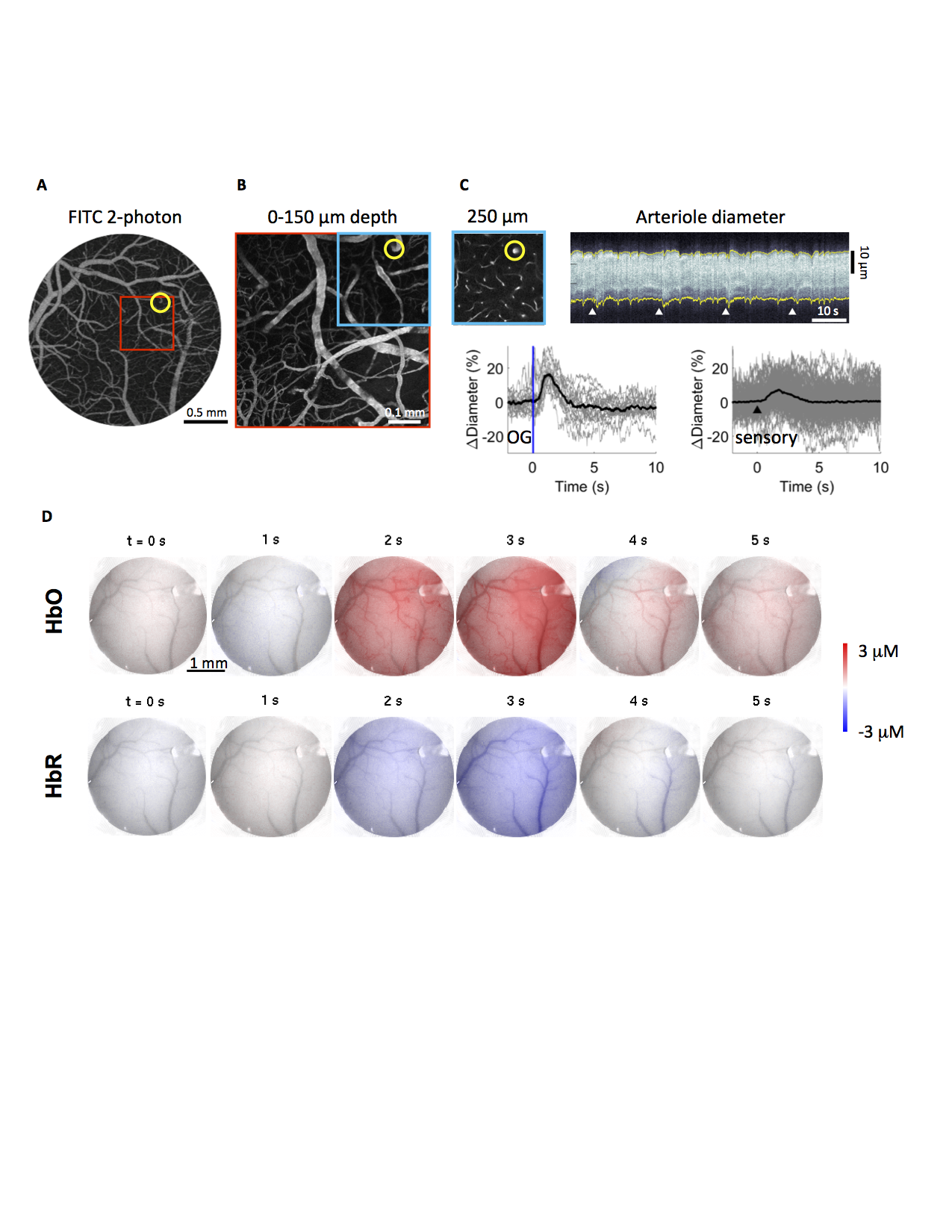
From two-photon microscopy to mesoscopic optical imaging providing a proxy for the BOLD signal.
A-B) Image of the surface vasculature at 4x (A) and 20x (B) zoom. C) A plane 250 um below the cortical surface; an example temporal diameter change profile acquired from the circled arteriole by repeated line-scans across the vessel. 4 white arrowheads indicate 4 stimuli onsets. Bottom: single-vessel dilation time-courses for optogenetic (N=19 trials) and sensory (N=160) stimuli. B) Spatiotemporal evolution of intrinsic optical signal changes (HbO, HbR) in a fully awake mouse in response to sensory stimulation.
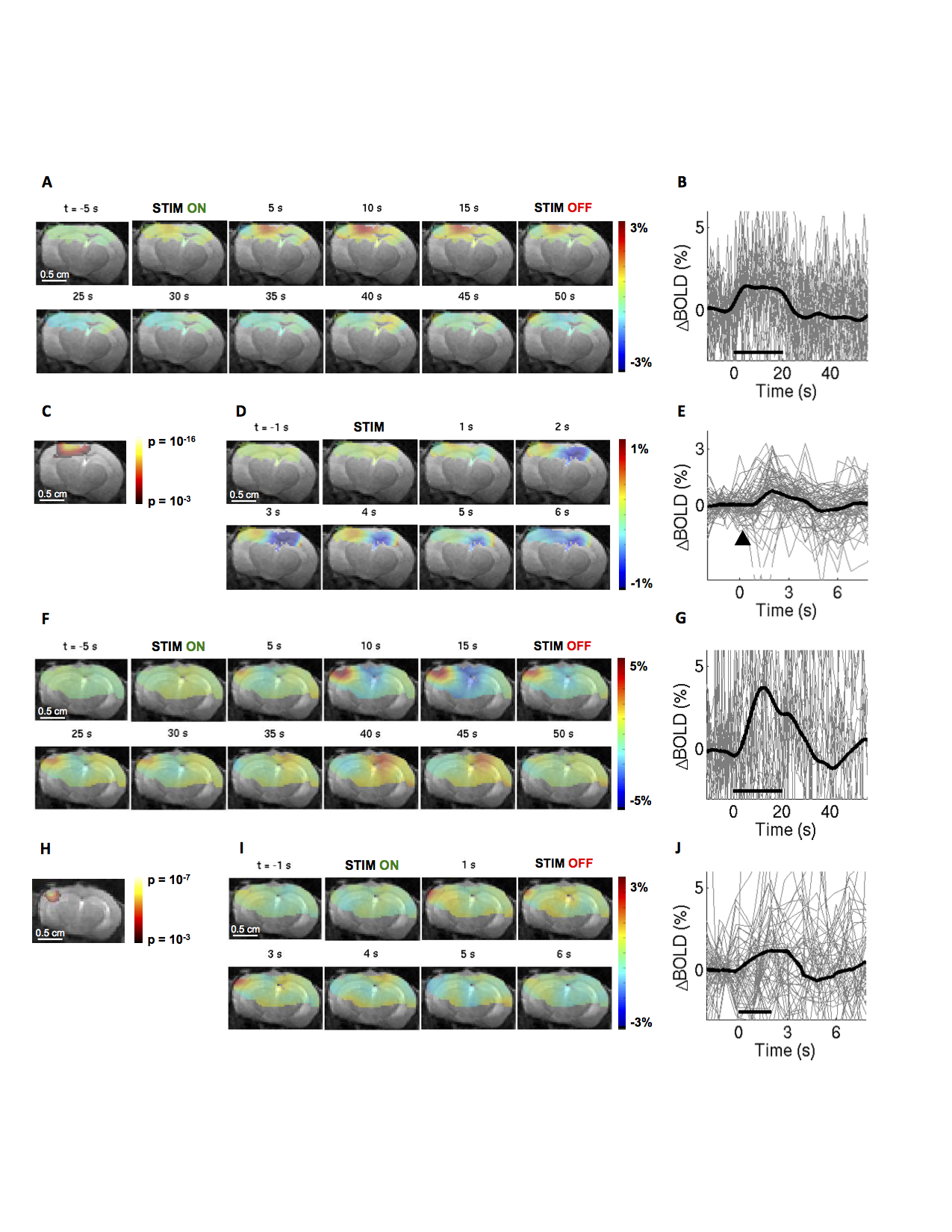
BOLD-fMRI response in a fully awake mouse.
A,D) Spatiotemporal BOLD response, in 1 slice presented as trial-averaged ratio maps overlaid on structural images, in response to a ''blocked'' (A) and ''event-related'' (D) optogenetic stimulation of excitatory neurons. EPI images were thresholded to reflect the sensitivity of the surface RF coil (for display purposes). B,E) BOLD response time-courses extracted from the active ROI. B) N=28 trials; E) N=69 trials; average overlaid in thick black. C) Thresholded statistical p-map assuming the standard hemodynamic response function (HRF) with temporal derivatives. F-J) As in A-E for sensory stimuli. A-C) N=21 trials. D-E) N=57 trials.
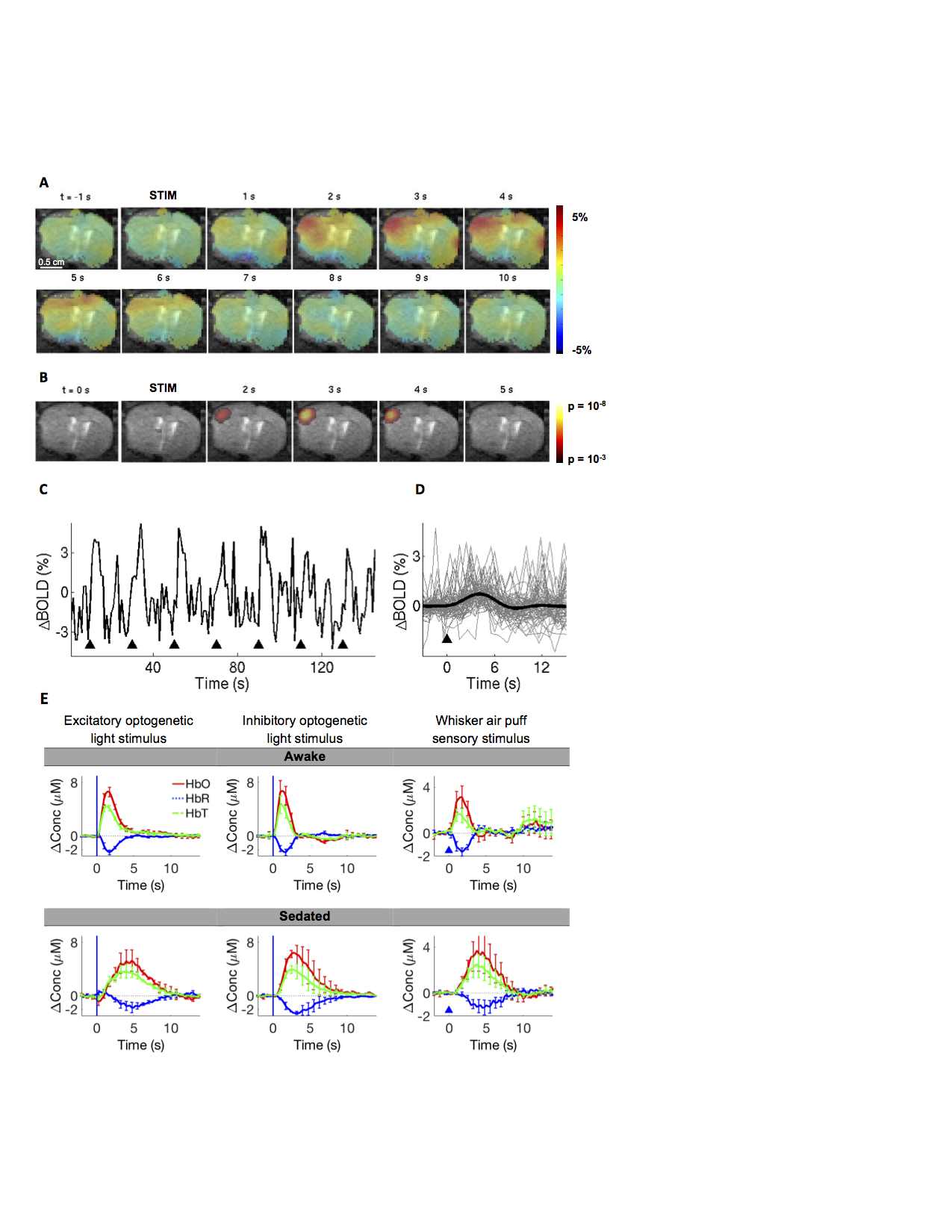
Effect of sedative drug on the hemodynamic response.
A) Spatiotemporal BOLD response to optogenetic stimulation in a sedated mouse (1.5 mg/kg chlorprothixene hydrochloride). B) Corresponding time-resolved p-maps, making no assumptions about the shape of the HRF (i.e. using a ‘’finite impulse response’’ model). C) ROI-averaged time-course. D) Trial-averaged, ROI-averaged timecourse (N=27 trials). E) Effect of sedation as measured using mesoscopic optical intrinsic signals imaging, providing a proxy for the BOLD signal. Three stimulus conditions are shown. Error bars indicate s.e.m. between subjects (n=3/2/5).