0202
Neural activity temporal pattern dictates the long-range brain-wide propagation pathways: An optogenetic fMRI study1Laboratory of Biomedical Imaging and Signal Processing, The University of Hong Kong, Hong Kong SAR, China, 2Department of Electrical and Electronic Engineering, The University of Hong Kong, Hong Kong SAR, China
Synopsis
The current overarching challenge in neuroscience is to establish an integrated understanding of the brain networks, particularly the spatiotemporal patterns of neural activities that give rise to functions and behavior. fMRI provides the most versatile neuroimaging platform for mapping large-scale activities in vivo. However, on its own, many important details of the underlying network activities remain unresolved. Here, we employed fMRI in combination with pulsed optogenetic stimulation paradigms to probe the spatiotemporal dynamics and functions of the well-defined topographically-organized somatosensory thalamo-cortical network. We reveal unique long-range propagation pathways that are dictated by distinct neural activity temporal patterns initiated from the thalamus.
Purpose
One grand challenge in contemporary neuroscience is to achieve an integrated understanding of the large-scale brain-wide interactions, particularly the spatiotemporal patterns of neural activity that give rise to functions and behavior1,2. Basic and clinical researchers have utilized fMRI to map local and large-scale brain functions via measuring bulk neuronal activities throughout the brain in response to sensory stimulation or cognitive tasks. However, fMRI only reflects the gross outcome of the complex and cascaded activities of various cell types and networks. One key missing piece is the ability to initiate specific neural activity patterns at a particular brain region and monitor their downstream propagation targets/pathways. In this study, we hypothesize that the temporal characteristics of neural activity train directly impact its downstream propagation pathway(s)/target(s). We examine the somatosensory thalamo-cortical network by employing pulsed optogenetic fMRI stimulation paradigms to activate ventral posteromedial thalamus (VPM) excitatory thalamocortical neurons. We aim to reveal the temporally-specific neural activity patterns that drive different activity propagation pathways within the well-defined topographically-organized somatosensory thalamo-cortical network3 and their functions.Methods
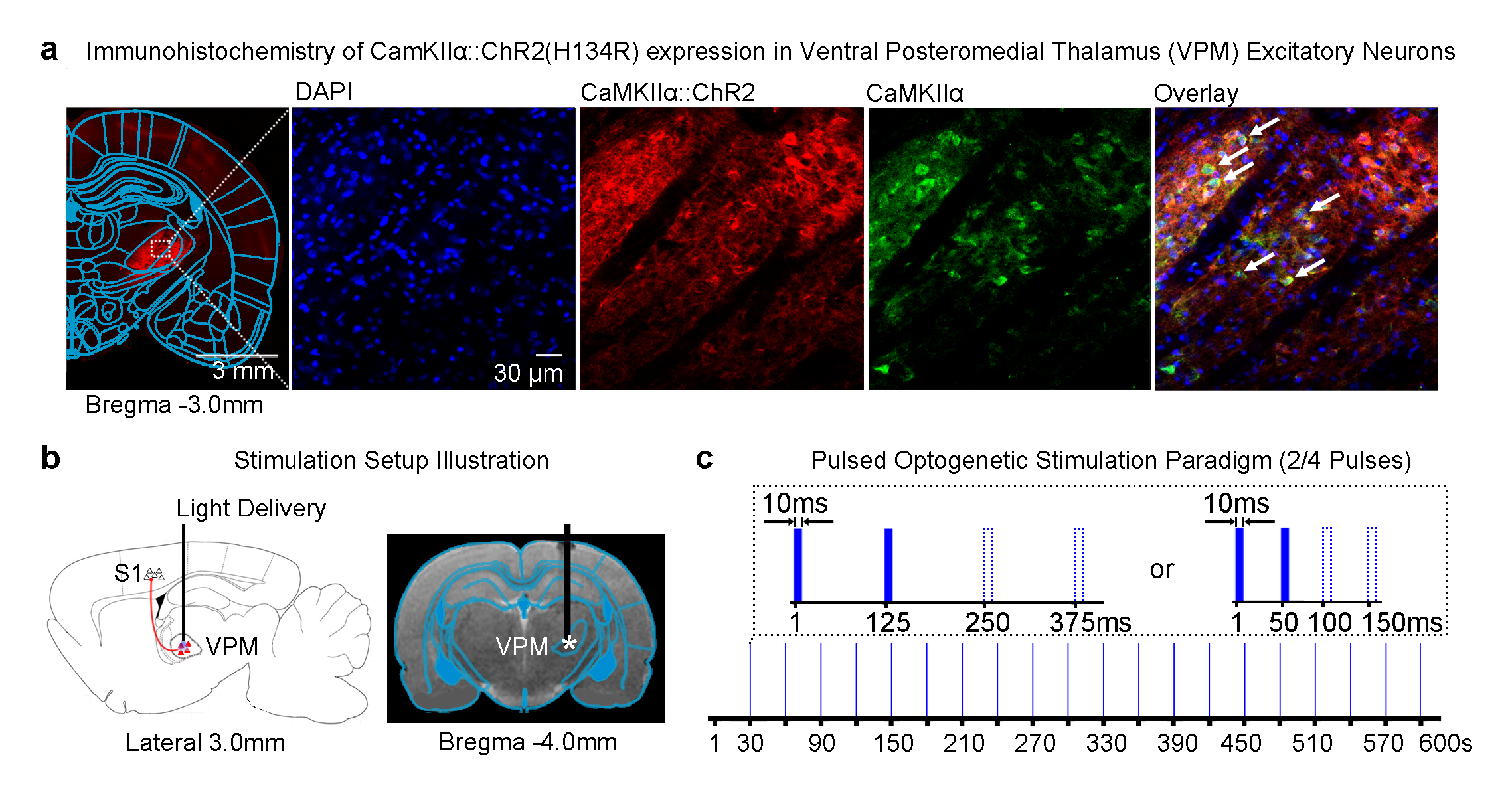
Optogenetics: 3μl of AAV5-CaMKIIα::ChR2(H134R)-mCherry was injected to right VPM of adult SD rats (200-250g, male, n=6; Figure 1a). Four weeks after injection, an opaque optical fiber cannula (d=450μm) was implanted at the injection site (Figure 1b). To characterize the brain-wide downstream propagation pathways, blue (473nm) light pulses were presented to ChR2 expressed in VPM (no. of pulses=2 or 4, interstimulus interval, ISI=125ms or 50ms, 10ms pulse width, 40mW/mm2; 20 stimulation blocks repeated every 30s; Figure 1c).
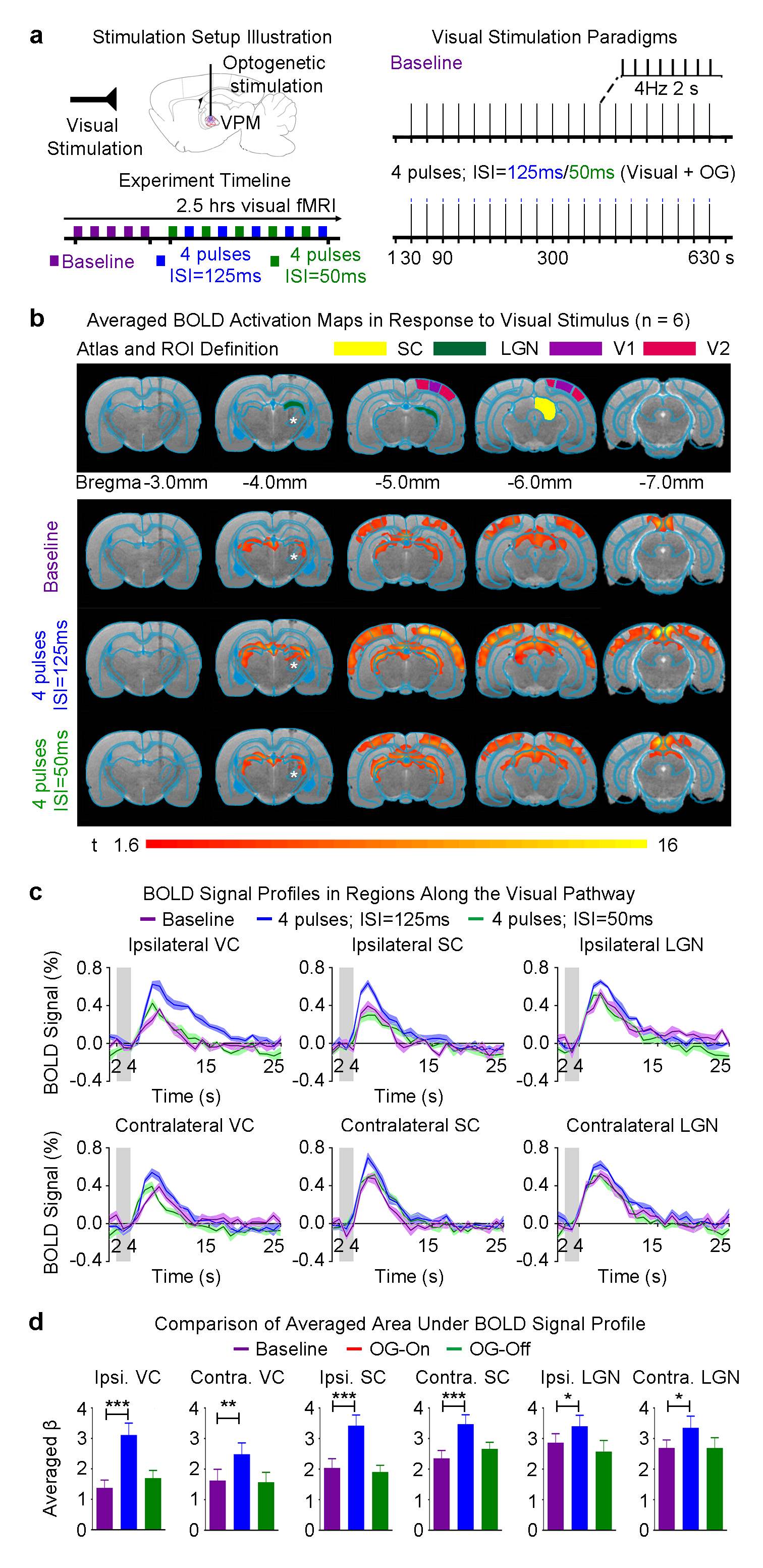
Visual stimulus with optogenetic stimulation: To examine the functional effects of optogenetic neuromodulation within somatosensory thalamo-cortical network, binocular visual stimulus (1Hz and 10% duty cycle; 0.5mW) was presented (30s light-off followed by 20 blocks of 2s light-on and 28s light-off) while blue light pulses were presented to right VPM (no. of pulses=2 or 4, ISI=125ms or 50ms, 10ms pulse width, 40mW/mm2).
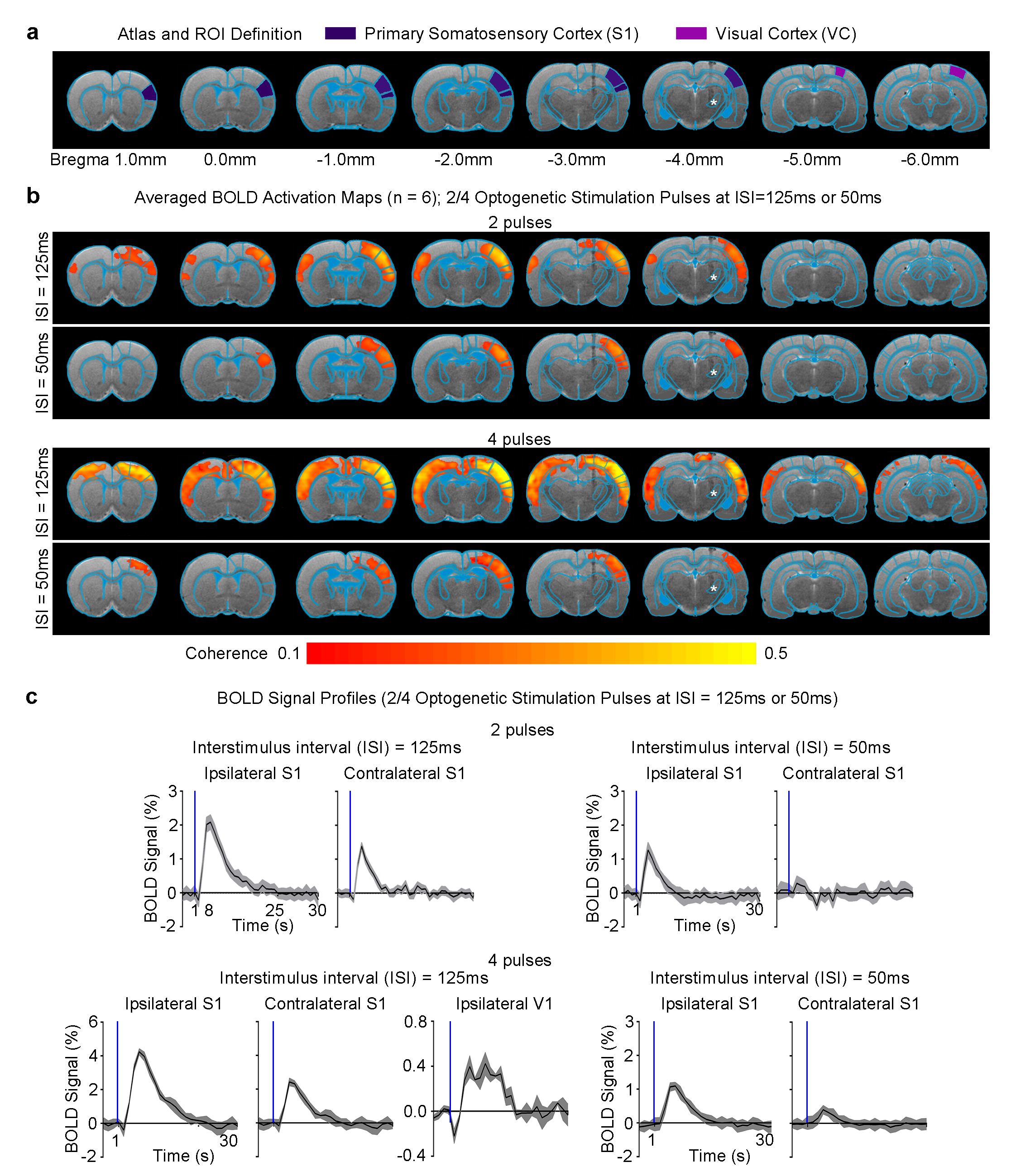
fMRI acquisition and analysis: fMRI data was acquired at 7T using GE-EPI (FOV=32×32mm2, matrix=64×64, α=56°, TE/TR=20/1000ms, 16 contiguous slices with 1mm thickness). Data were preprocessed before coherence analysis was applied to identify significant BOLD responses (P<0.001). BOLD signal profiles were extracted from anatomically defined ROI.
Results
Different VPM stimulation temporal patterns lead to different brain-wide neural activity propagation pathways across long-range projections in the thalamo-cortical network: All pulsed optogenetic stimulation at the VPM thalamocortical excitatory neurons activated the ipsilateral primary somatosensory cortex (S1; Figure 2). However, contralateral S1 was only activated when the ISI was 125ms, not 50ms, regardless of the number of stimulation pulses (i.e., 2/4). Interestingly, we found activations at bilateral visual cortex (VC) when the stimulation pulses were increased from 2 to 4 at an ISI of 125ms. Note that the BOLD response in ipsilateral VC was significantly stronger than that in the contralateral VC. Taken together, our findings demonstrate that different multiple long-range pathways (e.g., interhemispheric callosal and intrahemispheric cortico-cortical projections) were recruited when VPM thalamocortical neurons were excited with distinct optogenetic stimulation patterns.
Neural activity propagation from VPM to visual cortex enhances visual processing: As we found that bilateral VC were activated with 4 optogenetic stimulation pulses at 125ms ISI, we further examined the functional effects on visual processing. Baseline binocular visual stimulation as expected evoked positive BOLD responses bilaterally along the visual pathway, including the lateral geniculate nucleus (LGN), superior colliculus (SC) and VC (Figure 3b). In the presence of 4 pulses ipsilateral VPM optogenetic stimulation at 125ms ISI, visually-evoked BOLD responses significantly increased in bilateral VC, SC and LGN (Figure 3c). Overall, the visual responses in the ipsilateral brain hemisphere were enhanced more. However, visual processing was not affected when the optogenetic pulses were spaced at 50ms (Figure 3b, c).
Discussion and Conclusion
In this study, we revealed the distinct long-range neural activity propagation pathways/targets achieved by temporally-specific neural activity patterns initiated in the thalamus. We found robust positive BOLD responses in bilateral VC, in addition to bilateral S1, when the number of stimulation pulses was increased to 4 at 125ms interstimulus interval (ISI). However, BOLD response in contralateral S1 was absent when the ISI was decreased from 125ms to 50ms. These findings indicate that temporal characteristics of neural activity train plays a key role in determining the long-range propagation pathways/targets across long-range projections4,5. Furthermore, visual processing was enhanced when neural activity propagates to the VC. This observation indicates a possible neural mechanism underlying multisensory interactions between key sensory regions6,7. In summary, our work presents a novel and versatile optogenetic fMRI approach to investigate brain-wide neural activity propagation dynamics and functions.Acknowledgements
This work was supported by the Hong Kong Research Grant Council (C7048-16G and HKU17103015 to E.X.W.).References
1. Insel, T.R., Landis, S.C. & Collins, F.S. Research priorities. The NIH BRAIN Initiative. Science 340, 687-688 (2013).
2. Brain Research through Advancing Innovative Neurotechnologies (BRAIN) Work Group Report to the Advsiory Committee to the Director (US National Instutites of Health). (2014).
3. Diamond, M.E., von Heimendahl, M., Knutsen, P.M., Kleinfeld, D. & Ahissar, E. 'Where' and 'what' in the whisker sensorimotor system. Nat Rev Neurosci 9, 601-612 (2008).
4. Roland, P.E. Space-Time Dynamics of Membrane Currents Evolve to Shape Excitation, Spiking, and Inhibition in the Cortex at Small and Large Scales. Neuron 94, 934-942 (2017).
5. Jazayeri, M. & Afraz, A. Navigating the Neural Space in Search of the Neural Code. Neuron 93, 1003-1014 (2017).
6. Crevecoeur, F., Munoz, D.P. & Scott, S.H. Dynamic Multisensory Integration: Somatosensory Speed Trumps Visual Accuracy during Feedback Control. J Neurosci 36, 8598-8611 (2016).
7. Sieben, K., Roder, B. & Hanganu-Opatz, I.L. Oscillatory entrainment of primary somatosensory cortex encodes visual control of tactile processing. J Neurosci 33, 5736-5749 (2013).
8. Lee, J.H., Durand, R., Gradinaru, V., Zhang, F., Goshen, I., Kim, D.S., Fenno, L.E., Ramakrishnan, C. & Deisseroth, K. Global and local fMRI signals driven by neurons defined optogenetically by type and wiring. Nature 465, 788-792 (2010).
Figures