0173
The neurometabolic correlates of motor performance across the adult lifespan: a study on the effects of age1University of Leuven, Leuven, Belgium, 2icometrix, Leuven, Belgium, 3University of Leuven, leuven, Belgium, 4University Hospital Leuven, Leuven, Belgium
Synopsis
Aging is associated with alterations in neurochemistry of the brain, which can be assessed by MR spectroscopy. However, it’s unclear which metabolites are critical for function of the motor network. We explored whether changes in the neurometabolites of the aging brain account for motor-declines in bimanual coordination. We focused on neurochemistry of motor-occipital cortices as both regions are nodes of sensorimotor network underlying bimanual control. Myo-inositol was relevant for predicting Perdue test, which examine manual dexterity and general bimanual skills whereas N-acetylaspartate was associated with bimanual coordination task. Findings indicate NAA and mI could serve as biomarkers for integrity of motor network.
Introduction
Aging is associated with gradual alterations in structural and neurochemical characteristics of the brain, which can be assessed in vivo by using proton magnetic resonance spectroscopy (1H-MRS). This study aims to explore whether age-related changes in the metabolic profile of the aging brain account for motor performance declines associated with deficits in bimanual coordination. We focused specifically on the neurochemical integrity of the left sensorimotor cortex (SM1) and the occipital lobe (OCC) as both regions are thought to be main nodes of the sensorimotor network underlying bimanual control [1, 2]Methods
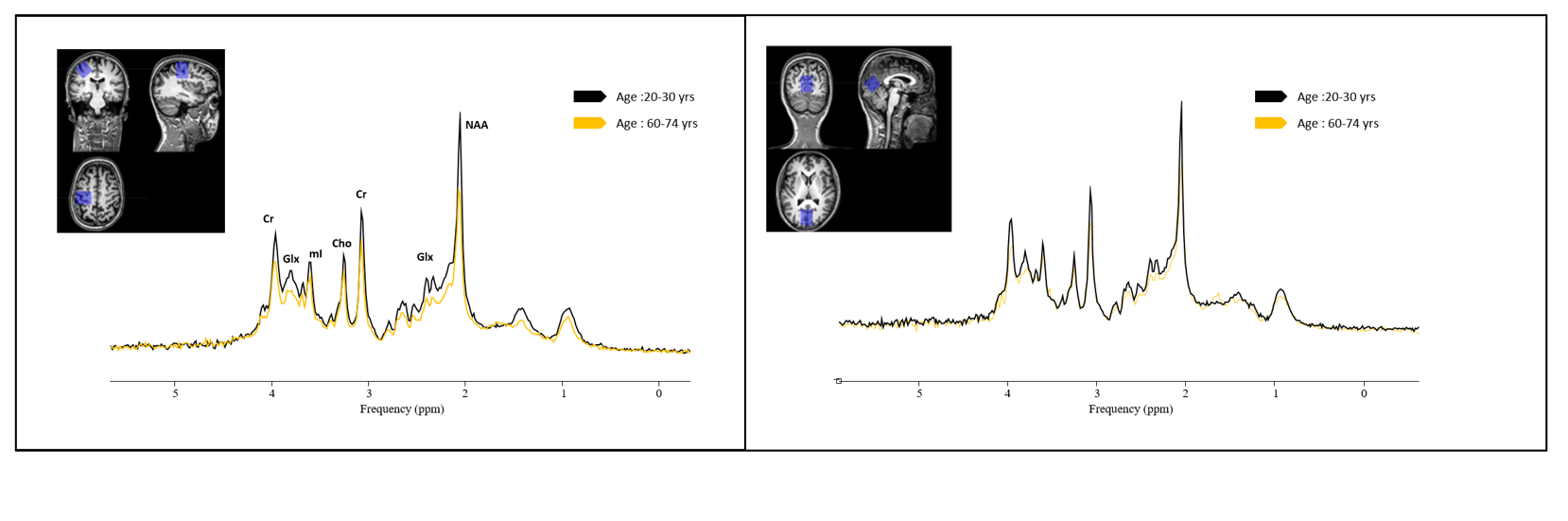
Single voxel 1H-MRS (PRESS, 3T Philips, TE/TR, 20ms/2s, NA = 128, 1.5x1.5x1.5 cm3 voxel) was performed in the SM1 and OCC regions of 106 healthy adults ((age range 20.0 – 74.5 years, 49 women). Previously described motor tasks, Purdue pegboard task (PPT) [3] and Bimanual coordination task (BCT)[4] were used in this study. MR spectra were processed and quantified using jMRUI v6.0 [5] and in-house developed software SPID [6] (Fig.1). N-acetylaspartate (NAA), creatine (Cr), choline (Cho), glutamate+glutamine (Glx), myo-inositol (mI), and taurine (Tau) were quantified. Tissue type correction was applied before quantification of metabolites[7,8]. Data for final analyses were obtained from 86 participants. Linear regression models were used to examine the effect of age on performance and neurometabolite levels. Potential relations between performance on PTT and BCT and estimated tissue-corrected metabolite concentrations in the two regions of interest were determined by calculating partial Pearson correlations, independent of the effect of age on performance. Stepwise multiple regression was used to determine the unique variance contributed by specific metabolitse to the performance of the PPT and BCT tasks.Results
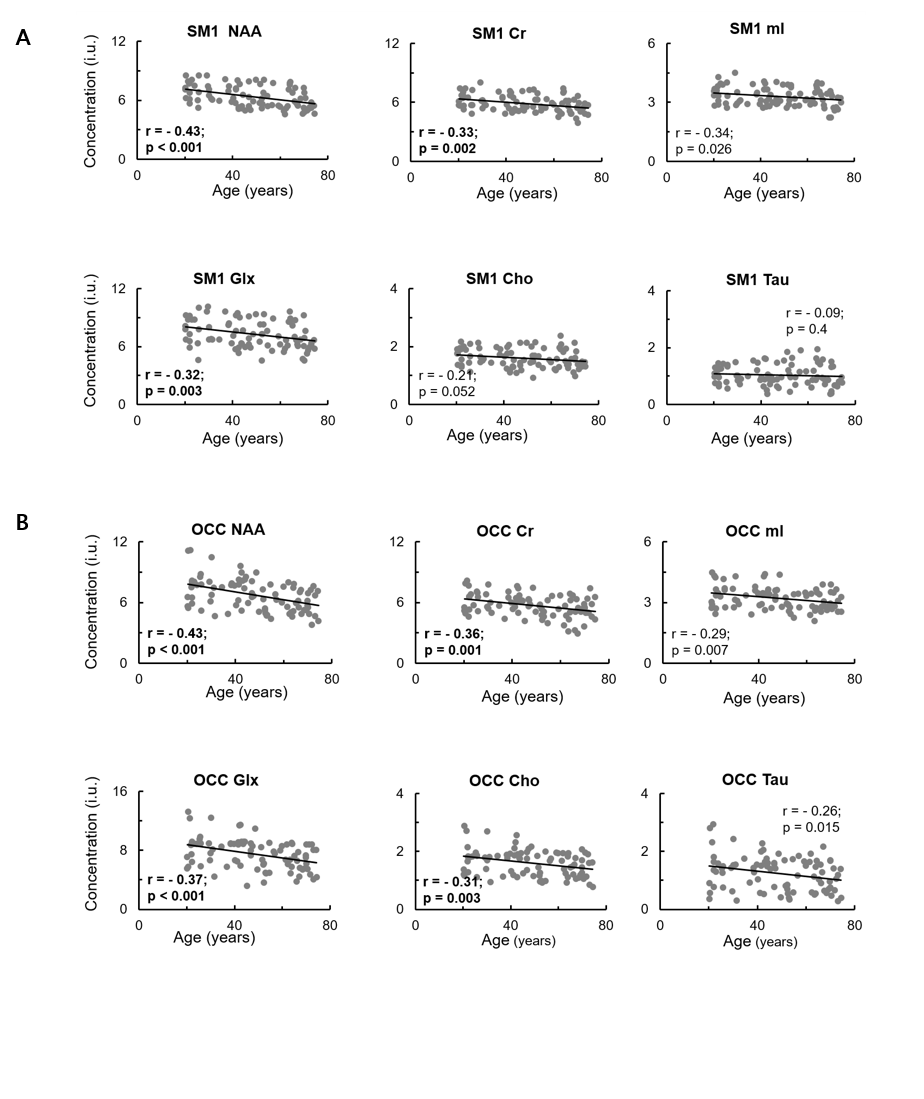
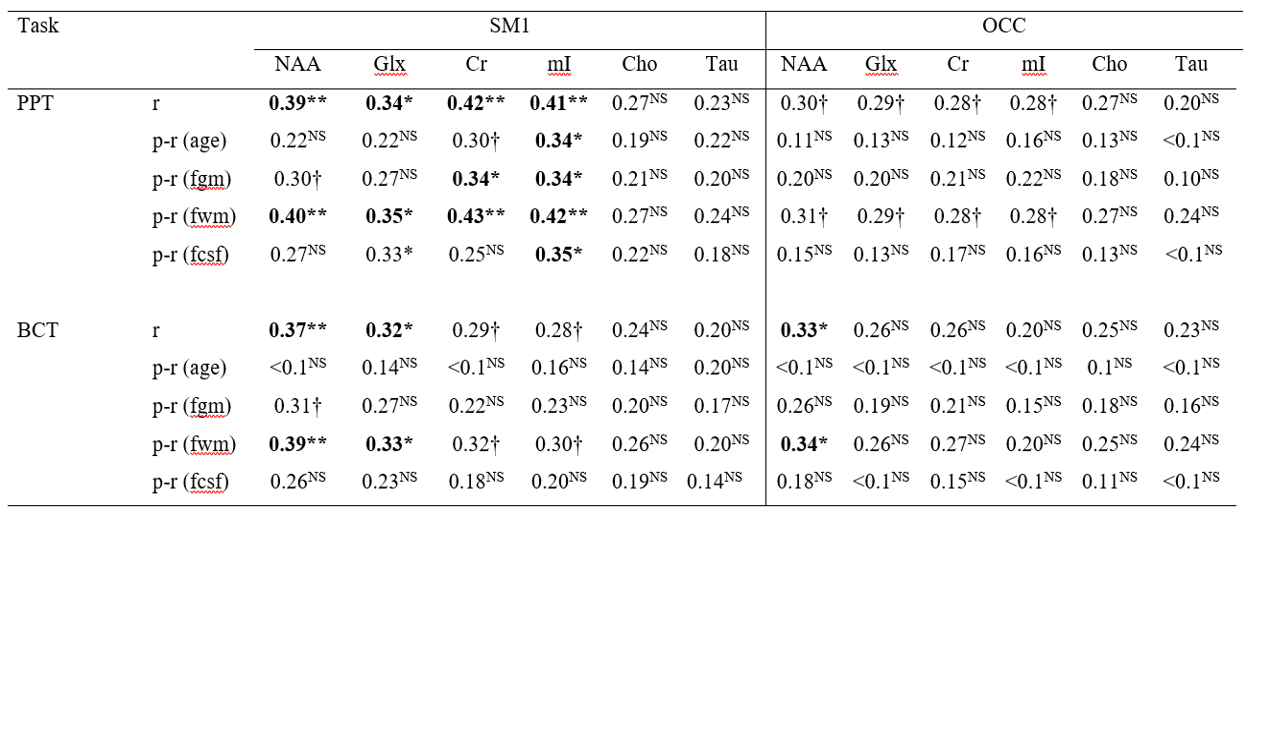
Significant negative correlations between age and brain metabolite concentrations in the SM1 were found for NAA (r = -0.43, uncorrected p < 0.001), Glx (r = -0.32, uncorrected p = 0.003), and Cr (r = -0.33, uncorrected p = 0.002) (Fig. 2A). Significant negative correlations between age and brain metabolite concentration levels in the OCC were found for NAA (r = -0.43, uncorrected p < 0.001), Glx (r = -0.37, uncorrected p < 0.001), Cr (r = -0.36, uncorrected p = 0.001), and Cho (r = -0.31, uncorrected p = 0.004) (Fig. 2B). Furthermore, data revealed significant positive associations between the average number of pairs inserted in the PPT and SM1 levels of NAA (r = 0.39, uncorrected p < 0.001), Glx (r = 0.34, uncorrected p = 0.001), Cr (r = 0.42, uncorrected p < 0.001), and mI (r = 0.41, uncorrected p < 0.001). For the BCT task, significant positive associations were observed between average accuracy scores on the four movement-trajectory conditions and SM1 levels of NAA (r = 0.37, uncorrected p < 0.001) and Glx (r = 0.32, uncorrected p = 0.002), and OCC levels of NAA (r = 0.33, uncorrected p = 0.002) (Table 1).Discussion
A major observation of the study was that tissue-corrected levels of multiple brain metabolite concentrations in both SM1 and OCC regions of the healthy human brain decreased significantly with age. This age-related decline in motor performance is potentially the consequence of low NAA and mI levels in the left SM1. Our findings provide evidence that changes in brain metabolite concentrations with aging in the left SM1 could account for age-related decline of bimanual coordination skills in healthy older adults. Low NAA levels in the left SM1 corresponded to poor performance on the BCT, a visoumotor task and with poor performance on the bimanual PPT, a task examining manual dexterity. Visually guided bimanual control was predicted by depletion in left-SM1 levels of mI.Conclusions
Our findings showed that aging of otherwise healthy subjects affects both, concentrations of brain metabolites and motor performance. Depending on the motor task, different metabolites have been identified as influencing factors of performance. Specifically, SM1 mI was found to be a relevant factor for predicting performance on the Perdue Pegboard test, which examins manual dexterity and general bimanual skills, whereas NAA was associated with performance changes on a complex bimanual coordination task. Taken together, our findings indicate that NAA and mI could serve as biomarkers for integrity of motor network supporting motor control in general and bimanual coordination in particular. Given that NAA and mI may reflect neurodegenerative processes related to alterations in WM microstructure, these findings highlight the dependence of bimanual control on the integrity of WM tracts within the SM1. Further research using 1H-MRS should be conducted to examine age-related changes in neurometabolite levels across multiple regions of the brain as plausible predictors of motor performance decline in healthy older adults.Acknowledgements
No acknowledgement found.References
1. King, B.R., van Ruitenbeek, P., Leunissen, I., Cuypers, K., Heise, K.-F., Santos Monteiro, T., Hermans, L., Levin, O., Albouy, G., Mantini, D., Swinnen, S.P., 2017. Age-Related Declines in Motor Performance are Associated With Decreased Segregation of Large-Scale Resting State Brain Networks. Cereb. Cortex 1–13.
2. Solesio-Jofre, E., Serbruyns, L., Woolley, D. G., Mantini, D., Beets, I. A., and Swinnen, S. P. (2014). Aging effects on the resting state motor network and interlimb coordination. Hum. Brain Mapp. 35, 3945–3961.
3. Serbruyns L., Gooijers J., Caeyenberghs K., Meesen R. L., Cuypers K., Sisti H. M., et al. (2015). Bimanual motor deficits in older adults predicted by diffusion tensor imaging metrics of corpus callosum subregions. Brain Struct. Funct. 220 273–290.
4. Sisti H. M., Geurts M., Clerckx R., Gooijers J., Coxon J. P., Heitger M. H., et al. (2011). Testing multiple coordination constraints with a novel bimanual visuomotor task. PLoS ONE 6:e2361910.1371/journal.pone.0023619.
5. Stefan, D., Cesare, F. Di, Andrasescu, A., Popa, E., Lazariev, A., Vescovo, E., Strbak, O., Williams, S., Starcuk, Z., Cabanas, M., Van Ormondt, D., Graveron-Demilly, D., 2009. Quantitation of magnetic resonance spectroscopy signals: The jMRUI software package. Meas. Sci. Technol. 20.
6. Poullet, J., Sima, D.M., Simonetti, A.W., De Neuter, B., Vanhamme, L., Lemmerling, P., Van Huffel, S., Neuter, B. De, Vanhamme, L., 2007. An automated quantitation of short echo time MRS spectra in an open source software environment: AQSES. NMR Biomed. 20, 493–504.
7. Gasparovic C, Song T, Devier D, Bockholt HJ, Caprihan A, Mullins PG, Posse S, Jung RE, Morrison LA. Use of tissue water as a concentration reference for proton spectroscopic imaging. Magn Reson Med. 2006; 55(6):1219-26.
8. Wansapura JP, Holland SK, Dunn RS, et al. NMR relaxation times in the human brain at 3.0 Tesla. J Magn Reson Imaging. 1999; 9:531–38.
Figures