0054
Exploration of the spatial and temporal characteristics of sensorimotor neural activity in the preterm human brain with simultaneous EEG-fMRI1Centre for the Developing Brain, Kings College London, London, United Kingdom, 2Department of Neuroscience Physiology and Pharmacology, University College London, London, United Kingdom, 3Department of Bioengineering, Imperial College London, London, United Kingdom
Synopsis
In the developing animal brain, different patterns of neural activity have distinct roles in the establishment of brain networks at different scales. Although studies suggest that the human preterm period is a crucial time for establishing brain connectivity, the role of different frequencies of neural activity has not been studied. We therefore used simultaneous EEG-fMRI and a robotic somatosensory stimulus to study the temporal and spatial characteristics of evoked neural activity in a group of preterm infants. Specific types of neural activity were associated with different BOLD responses, suggesting that these methods offer new insights into developing brain activity.
Introduction
In the neonatal animal brain, fast neuronal activity synchronizes local networks through a cortico-thalamic feedback loop, while slower oscillations propagate more widely across larger brain areas.1,2 It is thought that these different activity patterns play distinct roles in the formation of small and large scale cortical networks. In human preterm infants, EEG recordings are also characterized by a composite pattern of fast frequency ripples in the alpha-beta band (8-30Hz) coinciding with a high amplitude slow delta wave (0.5-4Hz), called delta brushes.3 These events occur spontaneously, but are also elicited by peripheral stimulation or sensory feedback from spontaneous motor behavior. We have previously demonstrated that simultaneous EEG-fMRI can be used to localize the source of spontaneous delta brushes.4 Here, we coupled these techniques with a precisely controlled somatosensory stimulus5 to characterize the cerebral correlates of evoked delta brush activity. We hypothesized that the different frequency components of these events would be related to the activation of networks with different scale.Methods
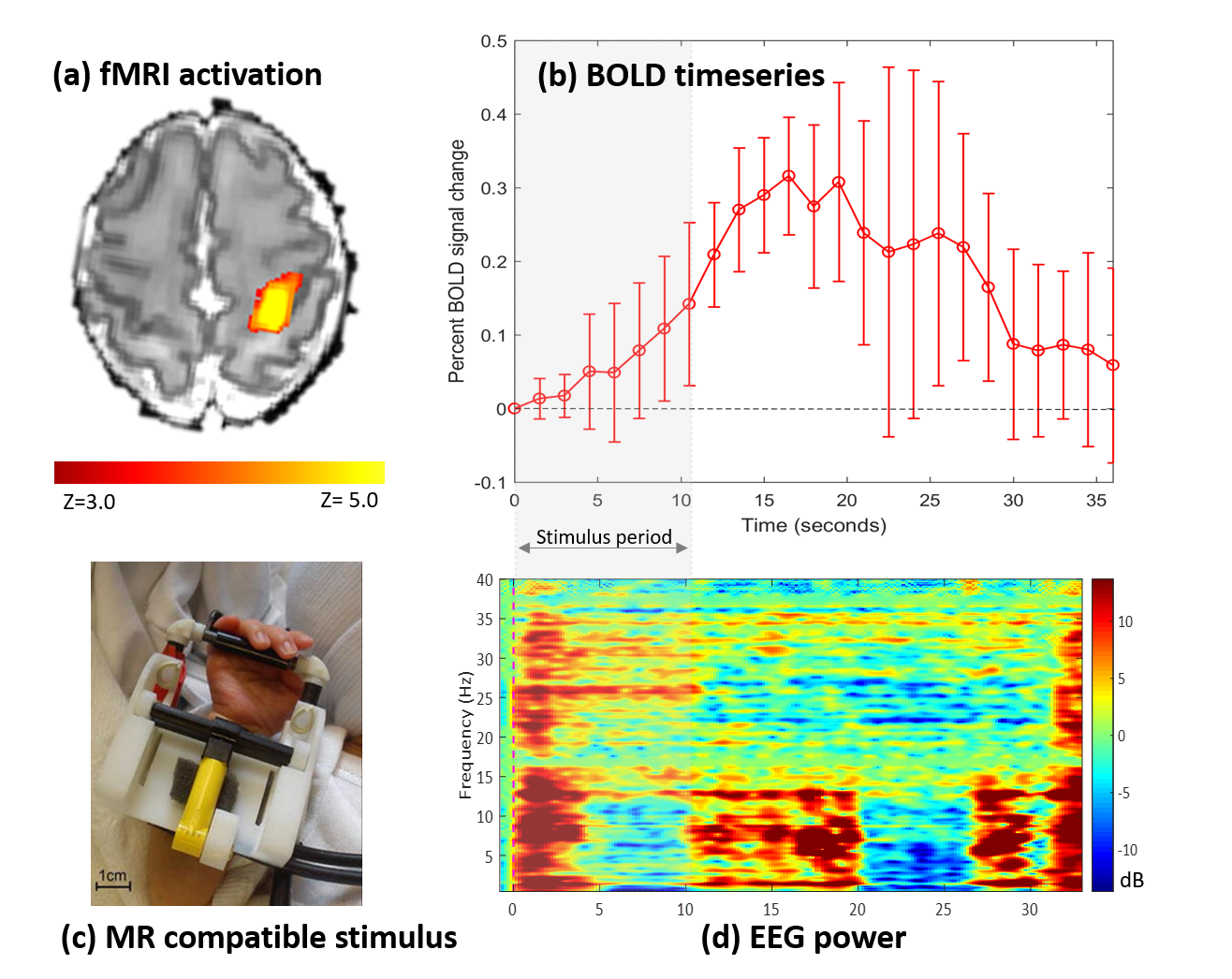
Simultaneous EEG-fMRI data was acquired during natural sleep from six preterm infants (post-menstrual age at scan 31+6 to 34+3 weeks) using a 3T MRI scanner and 32 channel head coil (Philips Achieva, Best NL) on the neonatal unit at St Thomas’ Hospital, London. Infants were fitted with a custom-made 25 electrode EEG cap (EasyCAP GmbH, DE) which was then connected to an MR compatible EEG system (Brain Products GmbH, DE). Somatosensory stimulation in the form of 1 Hz flexion/extension of the right wrist was delivered for 10.5s with a custom-built MR compatible robotic device.5 fMRI data were acquired with a T2*-weighted single shot gradient echo EPI sequence lasting 10 minutes with resolution: 2.5x2.5x3.3mm; TR: 1500ms; TE: 30ms; flip angle: 60 degrees; SENSE factor 2.
MR gradient artefact removal and a 40Hz low-pass filter were applied to the EEG data using Analyzer II software (BrainProducts GmbH, DE). Wavelet analysis was done in EEGLAB (Swartz Center for Computational Neuroscience ,USA). Spectral analysis of the peristimulus EEG was used to characterize temporal changes in frequency power (µV2) for the delta (0.5-4Hz) and alpha-beta (8-30Hz) bands recorded at contralateral electrodes C3 and CP3. In addition to a traditional box-car function derived from the timing of the block of stimulation, the spectral information was convolved with an age-specific HRF model to generate frequency-specific explanatory variables for general linear model analysis as implemented in FSL .6,7
Results
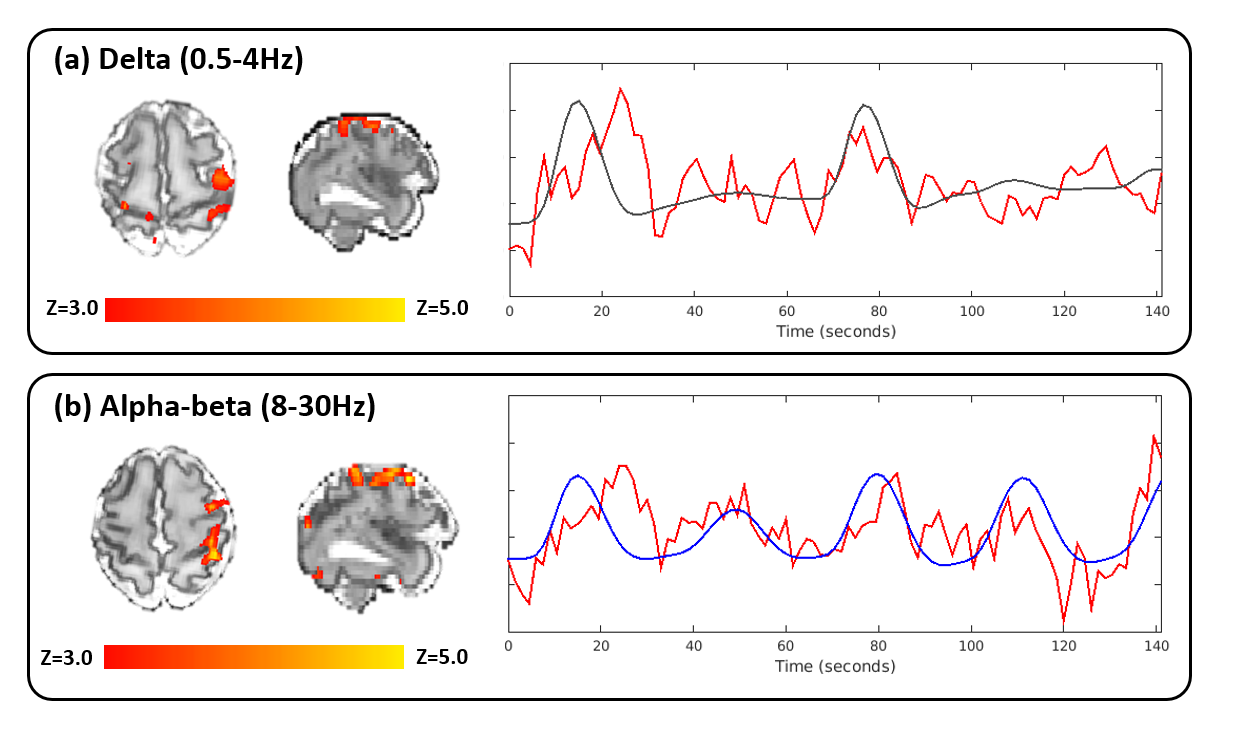
Data from one infant was excluded due to excessive head movement. In the five remaining infants, somatosensory stimulation of the right wrist evoked neural activity on the EEG which could be spatially localized with fMRI using the traditional box-car function to the contralateral (left) sensorimotor cortices (Figure 1). Time-frequency analyses of the EEG showed differential delta and alpha-beta power changes in response to both stimulus onset and offset. Significant clusters of BOLD activity were more commonly seen in association with temporal changes in alpha-beta power (5/5 infants) in comparison to delta power (3/5 infants). In infants with a significant BOLD response to both frequency bands, each band was associated with a different spatial pattern of BOLD response, suggesting that they may contribute differently to the identified hemodynamic responses (Figure 2).Discussion
This is the first study to utilize task-based simultaneous EEG-fMRI to identify the source of frequency specific evoked neuronal patterns in the immature human brain. In this initial dataset, somatosensory stimulation of the wrist was associated with a localized positive BOLD response in the contralateral primary sensorimotor cortex, and time-varying changes in delta and alpha-beta power in the contralateral central and centro-parietal electrodes on EEG. These distinct frequencies of neural activity may contribute differently to the induced BOLD response. Alpha-beta activity alone may induce a hemodynamic response in the preterm brain. In animals, these oscillations are likely due to glutamatergic currents originating from the thalamus and subplate,8 facilitated by the lack of feedforward inhibition because of the delayed maturation of cortical GABAergic circuits.9 This net excitation in cortical circuits is considered to be necessary for cortical map formation10 as its ceasing co-occurs with the end of a critical period of neuronal plasticity.11Conclusion
The combination of simultaneous EEG-fMRI and an automated robotic somatosensory stimulus can enable detailed characterization of both the temporal and spatial characteristics of induced neural activity in the developing human brain. In addition to providing new insights into maturation of the neurovascular coupling which underlies the BOLD response, these methods may also help to answer fundamental questions about how cortical specialization and connectivity emerge in early human life.Acknowledgements
The authors acknowledge support from the Medical Research Council (MRC) Clinician Scientist Fellowship (MR/P008712/1), MRC Career Development Award (MR/L019248/1) and from the Department of Health via the National Institute for Health Research (NIHR) comprehensive Biomedical Research Centre award to Guy’s and St Thomas’ NHS Foundation Trust in partnership with King’s College London and King’s College Hospital NHS Foundation Trust. We would also like to thank BrainProducts and EasyCap for technical support.
References
1. Yang J-W, Hanganu-Opatz IL, Sun J-J, Luhmann HJ. Three Patterns of Oscillatory Activity Differentially Synchronize Developing Neocortical Networks In Vivo. J Neurosci. 2009;29(28):9011-9025. doi:10.1523/JNEUROSCI.5646-08.2009
2. Yang J, An S, Sun J, et al. Thalamic Network Oscillations Synchronize Ontogenetic Columns in the Newborn Rat Barrel Cortex. 2013;(June):1299-1316. doi:10.1093/cercor/bhs103
3. Whitehead K, Pressler R, Fabrizi L. Characteristics and clinical significance of delta brushes in the EEG of premature infants. Clin Neurophysiol Pract. 2016;2:12-18. doi:10.1016/j.cnp.2016.11.002
4. Arichi T, Whitehead K, Barone G, et al. Localization of spontaneous bursting neuronal activity in the preterm human brain with simultaneous EEG-fMRI. Elife. 2017;6:1-14. doi:10.7554/eLife.27814
5. Allievi AG, Melendez-Calderon A, Arichi T, Edwards AD, Burdet E. An fMRI compatible wrist robotic interface to study brain development in neonates. Ann Biomed Eng. 2013;41(6):1181-1192. doi:10.1007/s10439-013-0782-x
6. Arichi T, Fagiolo G, Varela M, et al. Development of BOLD signal hemodynamic responses in the human brain. Neuroimage. 2012;63(2):663-673. doi:10.1016/j.neuroimage.2012.06.054
7. Jenkinson M, Beckmann CF, Behrens TEJ, Woolrich MW, Smith SM. Fsl. Neuroimage. 2012;62(2):782-790. doi:10.1016/j.neuroimage.2011.09.015
8. Dupont E, Hanganu IL, Kilb W, Hirsch S, Luhmann HJ. Rapid developmental switch in the mechanisms driving early cortical columnar networks. Nature. 2006;439(7072):79-83. doi:10.1038/nature04264
9. Daw MI, Ashby MC, Isaac JTR. Coordinated developmental recruitment of latent fast spiking interneurons in layer IV barrel cortex. Nat Neurosci. 2007;10(4):453-461. doi:10.1038/nn1866
10. Luhmann HJ, Khazipov R. Neuronal activity patterns in the developing barrel cortex. Neuroscience. 2018;368:256-267. doi:10.1016/j.neuroscience.2017.05.025
11. Fox K, Science N, Island R. A Critical Period for Experience-dependent Barrel Cortex Synaptic Plasticity in Rat. 1992;(May):1626-1636.
Figures