0036
Development-based Joint Parcellation of Human and Macaque Brains during Infancy1Department of Computer Science and Technology, Shandong University, Jinan, China, 2Department of Radiology and BRIC, University of North Carolina at Chapel Hill, Chapel Hill, NC, United States
Synopsis
Joint cortical parcellation maps of human and macaque infant brains with corresponding regions are highly desirable, since they provide basic common cortical parcels for both region-based and network-based studies of two closely-related species. Developmental patterns of infants indicate underlying rapid changes of microstructures, which determine the molecular and functional principles of the cortex. Leveraging 210 longitudinal human infant MRI scans and 140 longitudinal macaque infant MRI scans, we develop a novel method to generate the first joint parcellation maps of human and macaque infants based on their developmental patterns.
Introduction
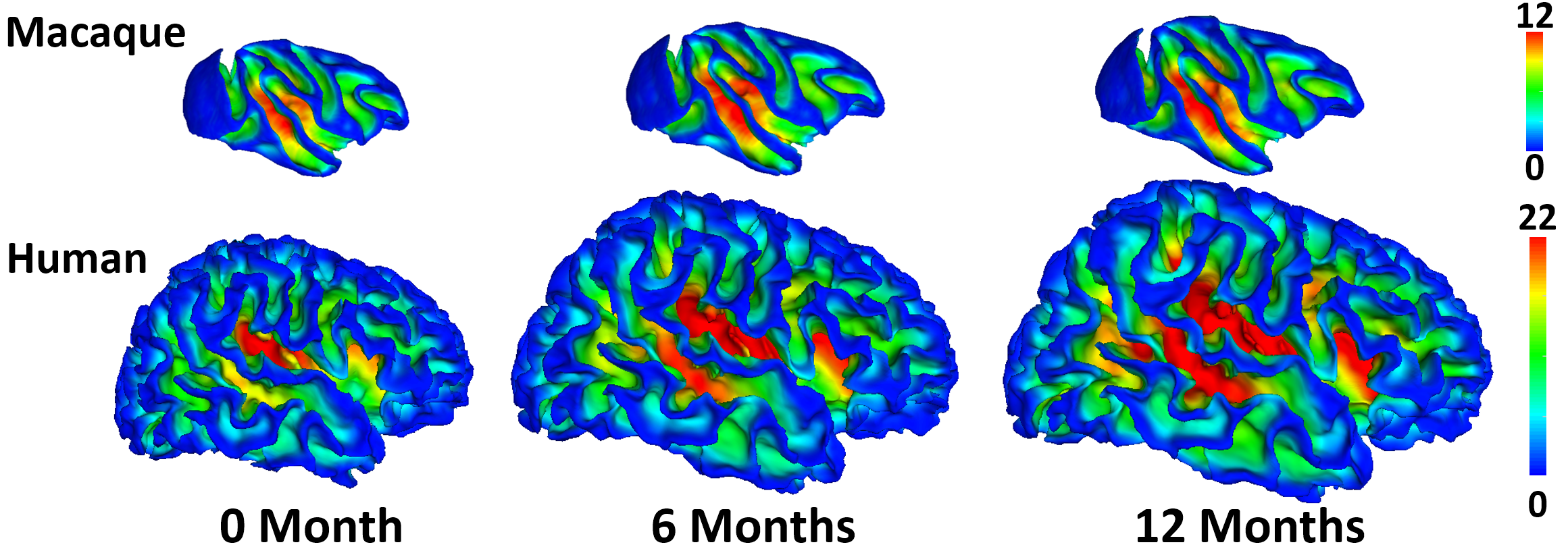
Comparative characterization of early postnatal brain development1,2 between humans and macaques using neuroimaging data plays an important role in understanding the underlying mechanisms of brain development, evolution, and neurodevelopmental disorders3-5. Parcellation aims to divide the cerebral cortex into a set of regions that are distinct in structure, function, or connectivity. Joint parcellation maps of human and macaque infant brains can thus provide common cortical basic parcels of the two closely-related primate species, thus enabling region-based and network-based comparisons in revealing mechanisms in brain evolution and development. However, due to huge differences in brain size and anatomical structures (e.g., cortical folding patterns) across species and age (Fig. 1), conventional parcellations typically based on anatomical landmarks are not suitable for defining the joint parcellation maps. Considering the dynamic development of both human and macaque infant brains, it is more meaningful to create joint cortical parcellation maps based on the developmental pattern of cortical properties (e.g., surface area), which indicate underlying rapid changes of microstructures and determine the molecular and functional principles of the cortex. Therefore, we unprecedentedly propose to jointly parcellate human and macaque infant cortical surfaces based on their developmental patterns.Methods
Longitudinal T1w and T2w human brain MRI scans were obtained from subjects enrolled in the Multi-visit Advanced Pediatric (MAP) Brain Imaging Study, with 36 normal infants (18 males) during the first two years. Each infant has 4 to 7 longitudinal scans, with the imaging acquisition parameters as in6. Longitudinal T1w and T2w macaque brain MRI scans were from a public neurodevelopment dataset, with 30 infant rhesus macaques (15 males) from 0 to 24 months7. Each macaque has 4 to 5 longitudinal scans with acquisition parameters detailed in7. We applied infant-dedicated tools for tissue segmentation, cortical surfaces reconstruction8, intra-subject surface registration9, within-species inter-subject surface registration. To establish inter-species cortical correspondences, we aligned two within-species mean cortical folding maps onto PALS-B12 human atlas and F99 macaque atlas, respectively. Finally, we resampled each surface and computed the surface area for each vertex. To comprehensively capture the characteristics of developmental patterns of cortical vertices, we define two feature vectors at each vertex, i.e., 1) growth trajectory and 2) growth correlation profile. Based on these two features, we construct two similarity matrices for each species. The first one is defined as the Pearson’s correlation of growth trajectories between each pair of vertices on the surface to capture “low-order” similarity of developmental patterns. The second one is defined as Pearson’s correlation of the growth correlation profile between each pair of vertices on surface to capture “high-order” similarity of developmental patterns. To effectively leverage both common and complementary information of human and macaque infants, we non-linearly10 fuse these four similarity matrices into a single matrix and then perform joint parcellation using spectral clustering11.Results
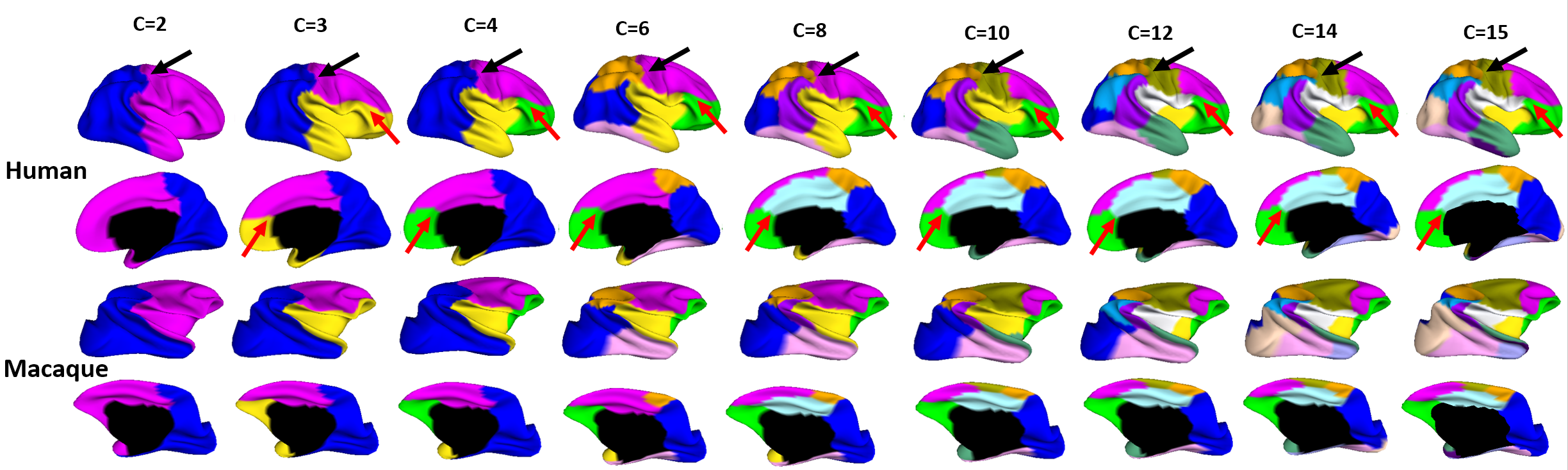
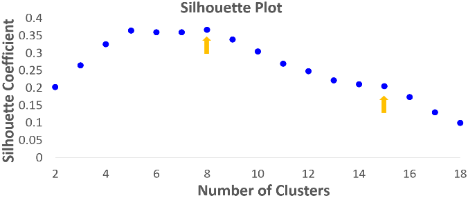
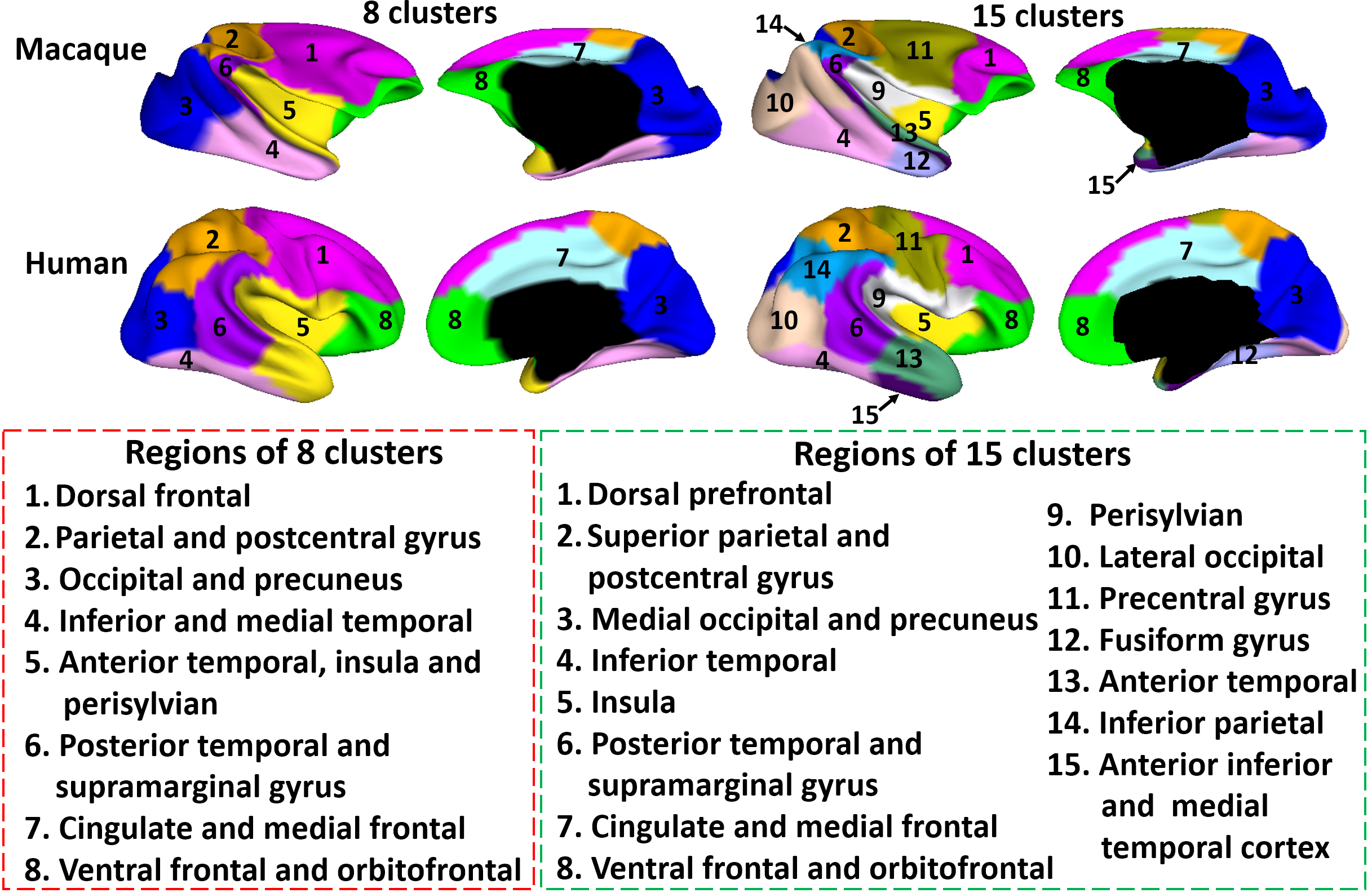
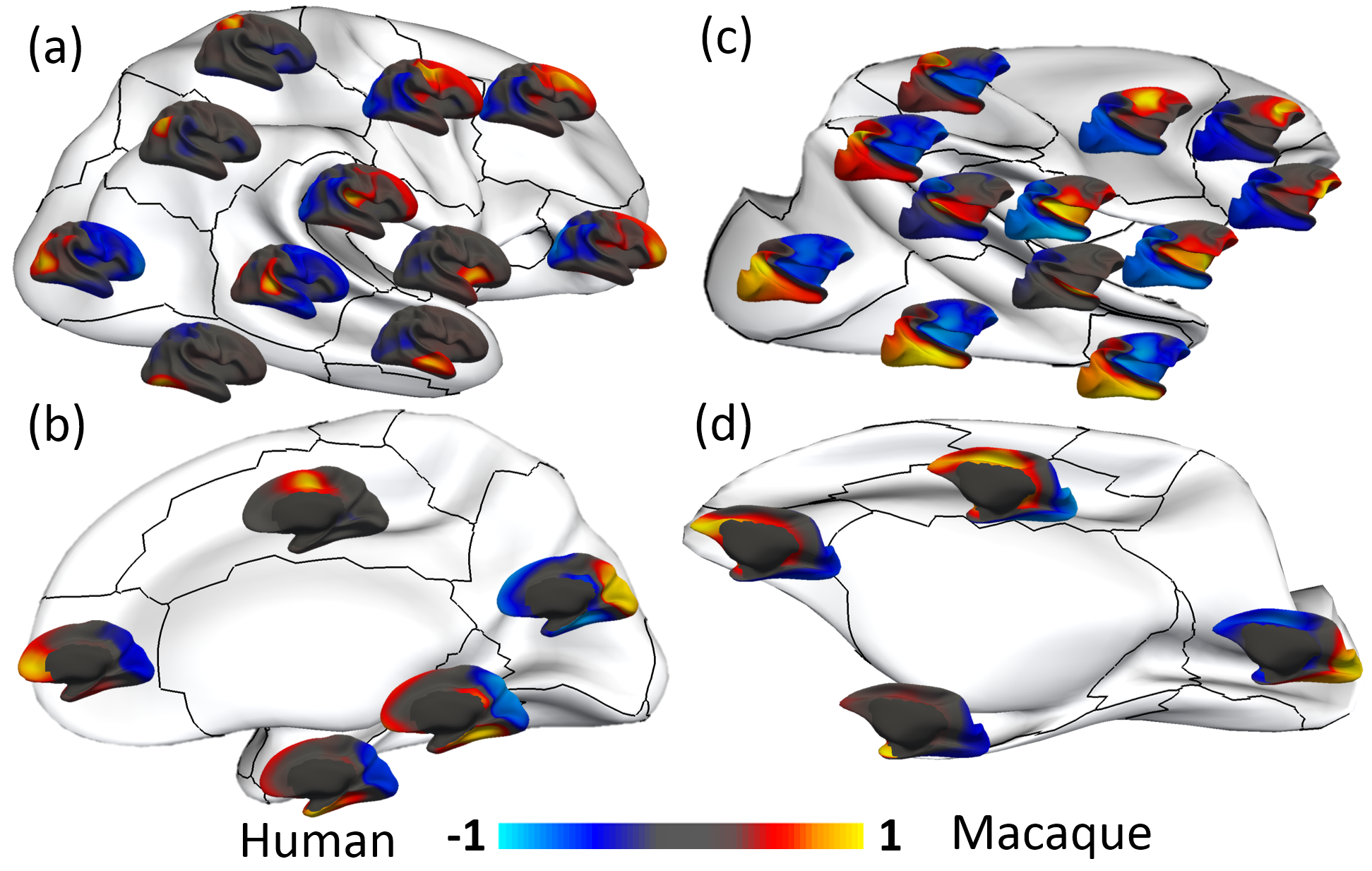
Fig. 2 shows the parcellation results with different cluster numbers from 2 to 15 using our proposed method, showing biologically-meaningful cortical regions. At 2-clusters parcellation, the proposed method identifies an anterior-posterior division. With the increase of the cluster numbers, the proposed method reveals a meaningful hierarchical organization of the growth patterning of surface area. For example, the boundaries between the frontal and parietal regions are well preserved from 2-clusters to 15-clusters by our method (as indicated by black arrows). The boundaries between the dorsal prefrontal and ventral prefrontal regions appear in 3-clusters, and are well preserved to 15-clusters by our method (as indicated by red arrows). Fig. 3 shows the silhouette coefficients of the joint parcellations by our method. The highest silhouette coefficient corresponds to 8-clusters. After that, the coefficient decreases and then reaches a relative stable plateau from 13-clusters to 15-clusters, and then decreases significantly after 15-clusters. Hence, we adopted 8-clusters and 15-clusters as our final results. As shown in Fig. 4, all clusters largely correspond to structurally and functionally meaningful specializations, with their approximated names shown in columns below the parcellation results. Fig. 5 shows the seed-based analysis at 15 clusters, which inspects the correlation patterns of growth trajectories of surface areas of humans and macaques, respectively. As shown in Fig. 5, in each species, the correlations between seeds and other vertices in the same cluster are always high, while the correlations between seeds and vertices in different clusters are low.
Discussion and Conclusion
We proposed a novel method for joint parcellation of human and macaque infant cortical surfaces based on their developmental patterns of cortical properties. Our charted development-based joint cortical parcellation maps reveal structurally and functionally meaningful specializations with a hierarchical organization.Acknowledgements
This work was partially supported by NIH grants (MH100217, MH107815, MH108914, MH109773, MH110274, MH116225, MH117943).References
1. Scott, J.A., et al., Longitudinal analysis of the developing rhesus monkey brain using magnetic resonance imaging: birth to adulthood. Brain Structure and Function, 2016. 221(5): p. 2847-2871.
2. Li, G., et al., Mapping region-specific longitudinal cortical surface expansion from birth to 2 years of age. Cerebral cortex, 2012. 23(11): p. 2724-2733.
3. Rilling, J.K., Comparative primate neuroimaging: insights into human brain evolution. Trends in cognitive sciences, 2014. 18(1): p. 46-55.
4. Hill, J., et al., Similar patterns of cortical expansion during human development and evolution. Proceedings of the National Academy of Sciences, 2010. 107(29): p. 13135-13140.
5. Van Essen, D.C., Cartography and connectomes. Neuron, 2013. 80(3): p. 775-790.
6. Li, G., et al., Construction of 4D high-definition cortical surface atlases of infants: Methods and applications. Medical image analysis, 2015. 25(1): p. 22-36.
7. Young, J.T., et al., The UNC-Wisconsin Rhesus Macaque Neurodevelopment Database: A Structural MRI and DTI Database of Early Postnatal Development. Frontiers in neuroscience, 2017. 11: p. 29.
8. Li, G., et al., Measuring the dynamic longitudinal cortex development in infants by reconstruction of temporally consistent cortical surfaces. Neuroimage, 2014. 90: p. 266-279.
9. Yeo, B.T., et al., Spherical demons: fast diffeomorphic landmark-free surface registration. IEEE transactions on medical imaging, 2010. 29(3): p. 650-668.
10. Wang, B., et al., Similarity network fusion for aggregating data types on a genomic scale. Nature methods, 2014. 11(3): p. 333-337.
11. Ng, A.Y., et al. On spectral clustering: Analysis and an algorithm. in Advances in neural information processing systems. 2002.
Figures